 Science
in Christian Perspective
Science
in Christian PerspectiveScience
in Christian Perspective
Evolution
Viral
Evolution:
Climbing Mount Molehill?
Ronald G. Larson*
2624 Pin Oak Dr.
rlarson@engin.umich.edu
Ann Arbor, MI 48103
From PSCF 52 (September 2000): 169.
Richard Dawkins' defense of Darwinism, which he describes as a climb up the "backside of 'Mount Improbable'," is challenged by direct observations of the evolution of RNA viruses, whose mutation rates are a million times faster than that of other organisms. These studies suggest severe limitations to gradualism, or "descent with modification" that Dawkins insists is an essentially complete explanation of origins. Comparisons of the genomes and biochemical machinery of viruses, as well as those of simple single-celled organisms, suggest that numerous lateral transfers of genetic material, or other non-Darwinian "saltations," are needed for a naturalistic explanation of origins.
Richard Dawkins and "Mount
Improbable"
The most articulate defender of Darwinism in modern times is
undoubtedly Richard Dawkins, the Charles Simonyi Professor of Public
Understanding of Science at Oxford University. His most important books are The
Blind Watchmaker1 and Climbing Mount
Improbable,2 which elegantly articulate the case for
gradual, Darwinian evolution of even the most intricate of nature's wonders,
such as the human eye or the web-spinning ability of spiders.
These examples are illuminated by engaging allegories, such as that of "Mount Improbable." On the front face of Mount Improbable is a sheer vertical cliff, impossible to climb, but on its back are "gently inclined grassy meadows, graded steadily and easily towards the distant uplands."3 The allegory represents his belief that miracles of biological "design" and "perfection" were achieved by a slow accumulation of small changes along a path of "continuous evolutionary gradation--a smooth incline up the mountain ..."4 The long march up this incline has transformed organisms with no eyes at all into ones containing superbly functional camera eyes, such as our own. It has also transformed primitive self-replicating molecules of the primeval "soup" into bacteria, multi-cellular organisms, and human beings.
This theory of biological change is none other than that of Charles Darwin, who argued that the processes producing all biological variation are occurring daily. These processes are random mutations combined with natural selection, the tendency of organisms whose biological endowment is ever so slightly better suited for survival than that of their parents to displace organisms less well-suited. In time, this gradual, but inexorable, process has produced the entire panoply of life, in all its resplendent complexity. Gradualism is the key to Darwinism. Darwin said it himself: "If it could be demonstrated that any complex organ existed which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down."5 Dawkins echos this key point as he discusses Darwinism: "There can be no sudden leaps upward--no precipitous increases in ordered complexity."6 That is, all biological change occurs by "descent with modification."
Dawkins applies Darwin's theory to the evolution of the human eye by expressing the theory as a series of simple queries. First he asks: "Could the human eye have arisen directly from something slightly different from itself, something that we may call X?"7 Given that the difference is slight enough, the answer Dawkins and most of us would give is "yes."
The next question follows: "Is there a continuous series of Xs connecting the modern human eye to a state with no eye at all?8 This would seem to be just a multiplication of the first question. Dawkins says: "The answer has to be yes, provided only that we allow ourselves a sufficiently large series of Xs ... Given, say, a hundred million Xs, we should be able to construct a plausible series of tiny gradations linking a human eye to just about anything!"9
Then comes the third question: "Considering each member of the series of hypothetical Xs connecting the human eye to no eye at all, is it plausible that every one of them was made available by random mutation of its predecessor?"10 He answers: "My feeling is that, provided the difference between neighbouring intermediates in our series leading to the eye is sufficiently small, the necessary mutations are almost bound to be forthcoming."11
Finally, the last question is: "Considering each member of the series of Xs connecting the human eye to no eye at all, is it plausible that every one of them worked sufficiently well that it assisted the survival and reproduction of the animals concerned?"12 Here is where many might answer "no." "What good is 5 per cent of an eye?" some ask. Or, considering the evolution of animal mimicry, wherein, e.g., an insect evades predators by looking uncannily like a twig, or, in another case, like a piece of dung, a scoffer may ask: "Can there be any edge in looking 5 per cent like a turd?" Dawkins does not flinch. He answers:
If I ... am walking through a forest at dusk, I may well fail to distinguish almost any dull-coloured insect from the twigs that abound everywhere. The image of the insect may pass over the edge of my retina rather than the more acute central region. The insect may be 50 yards away, and so make only a tiny image on my retina. The light may be so poor that I can hardly see anything at all anyway. The important thing about light intensity, distance of insect from predator, ... and similar variables, is that they are all continuous variables ... Such continuous variables foster continuous and gradual evolution.13
Dawkins rams home this point by drawing on his vast knowledge of biological diversity. His discussion of variations of the eye is particularly telling:
Some single-celled animals have a light-sensitive spot with a little pigment screen behind it. Among many-celled animals, various types of worm and some shellfish have a similar arrangement, but the pigment-backed light-sensitive cells are set in a little cup. This gives slightly better direction--finding capability ... In a continuous series from flat sheet of light-sensitive cells, through shallow cup to deep cup, each step in the series, however small the step, would be an optical improvement. Now if you make a cup very deep and turn the sides over, you eventually make a lensless pinhole camera ... The swimming mollusc Nautilus ... has a pair of pinhole cameras for eyes ... When you have a cup for an eye, almost any vaguely convex, vaguely transparent or even translucent material over its opening will constitute an improvement, because of its slight lens-like properties.14
Dawkins expands upon this explanation in his book, Climbing Mount Improbable, wherein the reader is taken on an excursion through the wide world of eyes: the cup eyes of the flatworm, the pinhole eyes of the Nautilus, the compound eye of the dragonfly, the "fibre-optic eye" of deep-sea crustaceans, the camera eye of humans, and others. By examining the distribution of eyes in the animal kingdom, it has been deduced that "eyes have evolved no fewer than forty times"; they seem to evolve at the "drop of a hat."15 In a computer model of eye evolution, Nilsson and Pelger started from a flat layer of photocells that could "evolve" randomly by small steps to permit both curvature and index of refraction variations to accumulate. Granting only a slightly higher survival probability to each slight improvement in the "eye," in only 364,000 generations, there appeared a computer version of a "good fish eye with a lens."16
Darwinism by Design
Dawkins provides an easy-to-understand computer simulation of the
principle of selection from random mutations using the example of a "monkey
typist." The monkey's first efforts on the typewriter produce the following
random string of characters:
WDLMNLT DTJBKWIRZREZLMQCO P
Dawkins now "breeds" from this incomprehensible starting point a litter of "progeny" in each one of which a letter is randomly changed to any other letter (with a space counting as a letter). Of all the offspring, only one is kept for continued breeding; the one whose letter sequence matches more closely, however, slightly, the Shakespearean phrase:
METHINKS IT IS LIKE A WEASEL
After forty-one generations of "breeding," the random initial phrase "evolves" into the target phrase: "METHINKS IT IS LIKE A WEASEL."17
This example illustrates the point that small random mutations, if selected upon nonrandomly, can accumulate to produce large changes. Also, in both the computer "eye" problem and the typing-monkey problem, the rate of evolution can be estimated from knowledge of the mutation rate and the specificity of the selection. The specificity of the selection, which is the degree of survival advantage gained from each small improvement, was chosen to be small in the simulation of "eye" evolution, and large in the typing-monkey problem. This explains the fast evolution of the random sentence of the typing monkeys in comparison to the computer-evolved "eye."
Dawkins has little patience with those who dismiss Darwinism as "a theory based on chance." He says Darwinism is "a theory of random mutation plus nonrandom cumulative natural selection."18 To Dawkins, the mutations are not important; they are random and ever present. What is important is that there is a "selection pressure," which favors mutations that push ever so slightly in a given direction, leading to accumulated change over time.
We can illustrate "selection pressure" by the thermodynamic principle that a gas confined by a piston in a cylinder, if heated slightly, will increase its pressure and then expand by pushing the piston out of the cylinder. The process by which this occurs involves tiny molecules striking the piston almost randomly from each side. However, because of the small temperature difference, the molecules inside the cylinder strike the piston slightly more energetically than those outside. If you look at only a few collisions, or even hundreds of them, you would not notice any particular difference in the force of the impact, and any difference that you might notice would be minuscule in comparison to the force needed to move the massive piston. Nevertheless, there are fantastically many collisions, so that even a slight bias in their intensity eventually produces a large effect. Although the motion of the piston might seem rapid to us, it actually occurs very slowly compared to the velocity of the molecules. We can explain this phenomenon without having a detailed theory of the dynamics of gas molecules; we only need to know that there is a bias in their energies of impact. Likewise, Darwin confidently predicted the occurrence of large evolutionary change, although he was completely ignorant of the mechanisms of mutation upon which his theory depended.
What
is important [to Dawkins] is a "selection pressure,"
which favors mutations
that push ever so slightly
in a given direction, leading to accumulated change over time.
The analogy of a sentence mutating by random changes in characters is apt in one sense at least: living things actually do have a kind of alphabet from which "sentences," "paragraphs," and whole "libraries" are constructed. The "letters" of the biological alphabet are the amino acids from which protein chains are built. Proteins make up the enzymes and other components of the machinery of life. There are twenty amino-acid "letters," rather than the twenty-seven letters of the "monkey" alphabet (counting the space as one letter). At the level of the DNA or RNA genetic code, a single amino-acid letter in the "alphabet" of life is coded for by a "codon," a triplet of three DNA bases.19 These bases consist of nucleotides given the letters A, T, C, and G. Thus, the triplet TAC codes for the amino acid tyrosine, which is given the letter "Y," while TTT represents the amino acid phenylalanine, which is given the letter "F." Since there are 43 = 64 different triplets that can be formed from the four-letter alphabet of the genome, while there are only twenty amino acids to be coded for, the genetic code has some redundancy. For example, the amino acid phenylalanine is coded for by both TTT and TTC.
Small changes in genetic information occur by mutations in the coding triplet. For example, a single copying error might change TAC to TTC, which would change the amino acid "Y" (tyrosine) into "F" (phenylalanine) in the protein synthesized by the cell. Such errors or "mutations" are known to occur, and their frequency has been measured. Such a mutation might produce no change in the functioning of the protein, a degradation in its functioning, or, much more rarely, a slight improvement in its functioning.
The
rapid mutation rate [of RNA viruses] implies that
they could in principle refresh their entire genome
in several thousand generations, which, thanks to their
short reproductive cycle time, is no more than a few years!
Fortunately, we know the time available for evolution, and the rate of mutation has been measured for various species. For eukaryotic life forms (those containing a cell nucleus), which include nonbacterial organisms ranging from yeast to humans, the mutation rate M is around 10-8 to 10-11 mutations per base pair per cellular replication cycle.20 (For complex multi-cellular organisms, the cellular replication time might be significantly less than that for replication of the entire organism.) Therefore, for eukaryotes, in particular multi-cellular ones, we can crudely estimate the cellular reproduction rate to be in the order of one year. Thus, a mutation rate of, e.g., 10-9 would imply that the organism could by random mutation refresh its entire genome in around 109 (or one billion) years, a period of time that corresponds nicely with the actual period of time available. (The age of the earth has been repeatedly measured at around 4.5 billion years.)
Thus, from this standpoint, the "Blind Watchmaker" hypothesis is reasonable. In fact, assuming a uniform rate of mutation, the genetic differences among organisms are sometimes used to estimate the approximate time period that has elapsed since two organisms diverged from their most recent common ancestor. This controversial dating method is based on the assumption of a molecular or genetic "clock" that ticks off mutations at a constant rate. (It is now known that different genetic "clocks" tick at very different rates, and perhaps at rates that vary with time, a finding that greatly complicates the use of such "clocks."21
However, to demonstrate empirically the truth of the "Blind Watchmaker" hypothesis, without having to resort to fictitious computer models, such as those for a "fish eye," or for the "typing monkeys," we need to examine a real biological system over a duration of time long enough to observe its steady evolution from one type of form of life into a radically different one. The time T required for an organism to completely refresh its genome is the cellular reproductive cycle time R, divided by the mutation rate M; that is, T = R/M. As noted above, however, this time T is in the order of a hundred million to a billion years for complex multicellular life forms. For simpler life forms like bacteria, the reproductive cycle time R can be reduced to a day or less, but, even so, T remains in the range of millions of years. For a human, a mutation in a single gene containing, e.g., 1000 bases, might take thousands of generations. Therefore, Dawkins justifies the use of crude computer models to back up his evolutionary arguments. He says: "Humans haven't the patience to wait a million generations for a mutation!"22
Viral Evolution: A Modest
Proposal for the "Blind Watchmaker"
Amazingly, however, there exist microbes for which the time T is
experimentally accessible! These are the RNA viruses. A virus is an entity that
is simpler than the simplest free living cell. It cannot generate energy or
reproduce on its own; it reproduces by infecting a living cell and hijacking
that cell's reproductive machinery, forcing it to manufacture viral components,
rather than those of the cell. In addition to their short reproduction time (a
few hours or less), RNA viruses have an extraordinarily high mutation rate M,
around 10-3 to 10-5!23
The reason for the high mutation rate is that the elaborate error-checking and repairing mechanisms that cells and DNA viruses employ during the replication of DNA do not operate in the replication of RNA. The high error rate in RNA replication may actually be beneficial to the RNA virus, since it can then mutate and evolve rapidly in response to attacks by the organism's immune system, or by anti-viral drugs administered to combat the virus. RNA viruses, such as the influenza virus, are notorious for their agility in evolving resistance to vaccines and anti-viral drugs. Dimmock and Primrose say: "An RNA virus can achieve in one generation the degree of genetic variation which would take an equivalent DNA genome 300,000 generations to achieve."24
For purposes of studying evolution, therefore, RNA viruses are ideal. Their rapid mutation rate implies that they could in principle refresh their entire genome in several thousand generations, which, thanks to their short reproductive cycle time, is no more than a few years! In addition, RNA viruses have genomes of modest size. In some cases, only around ten thousand bases, or "letters" of the genetic code, are enough to specify the entire virus. While this many letters might be analogous to, e.g., a whole chapter of reading material, it is a far cry from the whole library of genetic material present in the three-billion-base genome of a human. Thus, comparisons of the evolutionary family tree of viruses should be readily possible at a level of detail that extends down to that of individual bases. One might even hope eventually to reproduce biochemically the "stepping stones" by which virus "A" might have evolved into a completely different virus "B" by creating in the laboratory "missing-link" viruses that have attributes intermediate to those of "A" and "B." Thus, viral evolution is an area ripe for testing evolutionary hypotheses. Domingo and Holland, for example, note:
Limited complexity of genomes ..., their high mutability, rapid replication rate, and the possibility of testing the effects of several millions-fold differences in the population numbers of replicating genomes, confer great value upon viruses as model systems for understanding molecular evolution.25
While work on RNA viral evolution is in its infancy, early work has already yielded results that bear directly on the "Blind Watchmaker" hypothesis. Remember that, as stated by Dawkins, this hypothesis asserts that when a self-replicating system faces a fitness hurdle to its survivability, it will evolve novel and unanticipated functions. In the case of RNA viruses, experimental versions of the monkeys with typewriters have recently been realized. In these experiments, viruses are allowed to reproduce in a cellular culture; progeny viruses are harvested, and a randomly-chosen fraction of the resulting viruses are then used to infect a fresh cellular culture. Changes in the "fitness" of a viral population from generation to generation are directly measured by its infectiousness (that is, the number of progeny produced) relative to a reference virus grown under the same conditions.26
What do these experiments show? They
show that, starting from an initial "wild-type" virus, viral fitness
improves from generation to generation in just the way Darwin predicted. Fig. 1
shows that the fitness of vesicular stomatitis virus (VSV) improves
exponentially (that is, linearly on a semi-log plot) during multiple passages
through a cell culture, up to around one hundred passages. Thus, ten-fold
improvements in fitness can actually be observed in the laboratory, thanks to
the fast reproduction and high mutation rate of RNA viruses.27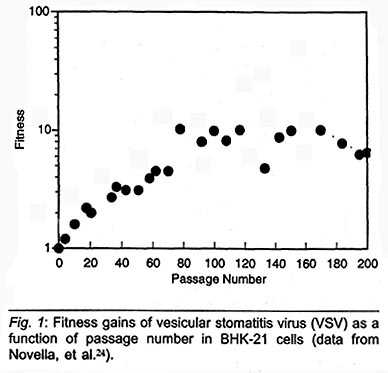
While some viruses improve their fitness, as we observe in Fig. 1, others actually become less fit over time.29 How could this happen? The experiments in which the fitness decreases with time are ones in which only a few, or even one virus particle is taken from a reproducing colony to form the ancestry of the next-generation colony. This highly constricted survival rate is imposed to mimic the effect of viral transmission from one host to another, which might occur by way of only a few, or even only a single viral particle.30 The reason that fitness can decrease under such constrained conditions of reproduction is now understood. It is due to the overwhelmingly high ratio of unfavorable mutations to favorable ones. When the viral colony faces a mass slaughter that leaves, e.g., only a single survivor, then the rare superior virus present in the colony is most likely to have been cut down along with the rest. Thus, the single virus that remains to produce offspring owes its survival, not to superior genes, but to luck. In all probability, the survivor will be an inferior representative of his formerly numerous colony. In this case, the following proverb holds good:
"The race is not to the swift; nor the battle to the strong ... but time and chance happeneth to them all" --(Eccles. 9:11).
This phenomenon, whereby a colony of organisms loses fitness due to "bottlenecks," or constrictions in the number of surviving descendants, is known as "Muller's Ratchet.31" The rare superior virus can break the ratchet only in prosperous times, during which it can combine ample resources with its superior growth and reproductive capabilities to gradually outproduce and overwhelm the vast hordes of inferior rivals.
Figure 1 shows the development of viral fitness under "prosperous" conditions in which VSV particles were transferred from one culture to the next. As we noted, fitness improves exponentially for around one hundred passages. After that, fitness levels off, and no steady gains in fitness are observed, only fluctuations in fitness. These recent results show that even under favorable conditions there is an apparent limit to viral fitness, at least within a given host medium over the duration of time tested. Thus, time plus the availability of mutations is not by itself a sufficient condition for continuous evolution.
Time
plus the availability of mutations
is not by itself
a sufficient condition
for continuous evolution.
Although these experiments are limited in their numbers of reproductive cycles, their results are consistent with other observations. The range of mutations actually permitted to a given virus has been observed to be limited, over the time scale for which direct observations are made (which, as noted, is as long or longer than the time T required to "refresh" the genome). Thus, experiments with picornaviruses (a class that includes the common cold) show frequent recurrence of the same subsets of mutations. Domingo and Holland note:
Even during several decades of viral evolution, variable positions in the capsid [coat] proteins of picornaviruses are alternatively occupied by a small subset of all possible amino acids, and a true accumulation of amino acid substitutions is not observed.32
Many virus codons and the corresponding amino acids are highly conserved. These include regions of the RNA associated with gene regulation, and even some receptor recognition sites where the virus is sensitive to antibody attack.33 Mutations at these and other positions on the RNA strand are not tolerated. As a result, the range of genetic variation available to each viral type is narrow. Beyond a certain number of replication cycles, the drift of the viral genome becomes limited as the virus explores new territory less frequently and instead revisits its old haunts again and again.
What can we make of reports of the emergence of "new" viruses? Periodic outbreaks of apparently new viral diseases are recurrent in human history; the AIDS epidemic is only a recent example. New strains of the influenza virus emerge episodically, including the disastrous 1918 "Spanish influenza" pandemic that killed twenty million people worldwide. However, instances in which the origins of a "new" disease have been tracked down suggest that in each case the virus responsible for the outbreak is usually not really new, but is instead a transplant from another species. For example, it now appears that new influenza strains, which frequently emerge from China, are carried from waterfowl to pigs, and then to humans. This "viral traffic" is stimulated by integrated pig-duck farming common in China.34
Many other examples are now available in which viral outbreaks are traced to inter-species migration of an existing virus, often because of a change in agricultural or other practices. In the case of the H2N2 influenza strain, for which an outbreak occurred in 1957, blood sera taken from people who had been alive in 1889 showed the presence of these same H2N2 antibodies, demonstrating that this "new" virus had merely been "hibernating," perhaps in an animal vector. After the earlier, resistant population had largely been replaced by a vulnerable younger population, H2N2 was able to re-emerge with renewed virulence. Morse summarizes: "Over the period of recorded history, emerging viruses have usually not been newly evolved viruses. Rather, they are existing viruses conquering new territory.35"
Thus, large changes in viruses seem to require mechanisms other than the accumulation of single- base "point" mutations, the kind represented by the typing monkeys. One such additional mechanism might be the evolution of the host cell. This might provide a change in viral environment, and open up the range of permitted mutations in a new direction. Another possibility is genetic re-assortment or recombination. The genome of influenza A virus, e.g., consists of eight single-strand RNA segments, each coding for one or two proteins. If a cell is simultaneously infected with two strains of this virus, the RNA segments of the two are randomly assembled in the next-generation virus. These hybrid viruses are genetically stable and can be produced in both cell cultures and whole animals. A hybrid strain of influenza recently infected harbor seals off the coast of Maine, killing 20% of the population.36
A fascinating case in which a somewhat novel virus may have emerged through recombination is that of the Western equine encephalitis virus (WEE), a type of togavirus. Togaviruses receive their name from the distinctive lipid bilayer these viruses wear over their protein coats like a Roman toga. After the genome of this virus was sequenced, it was found that the two glycoproteins of WEE are very similar to those of the Sindbis virus, while the portions of the genome coding for replication enzymes and the coat protein are similar to those of the Eastern equine encephalitis (EEE) virus.37 The simplest explanation for this is that WEE arose from recombination of genetic components borrowed from both Sindbis and EEE "parents." When this may have occurred is not known precisely, but based on the sequence differences and rates of mutation for this virus, it seems to have occurred at least one hundred years ago, and maybe much earlier.38
However, the change produced by this apparent recombination represents a rather minor degree of evolution. WEE, EEE, and Sindbis are all alphaviruses, which are mosquito-borne members of a single genus within the family of togaviruses. All three viruses have genomes of similar size and structure that are segmented identically and replicate themselves similarly. The modular arrangement of the genomes of viruses creates many possibilities for genetic "mixing and matching," whereby modular genetic units are swapped. Thus, when gene re-assortment occurs (which can be very rare or perhaps nonexistent in some viruses),39 it usually produces relatively minor changes, at least when compared to the changes required for the appearance of altogether new viral forms.
In fact, there is an enormous diversity of viral forms. Viruses differ one from another in their genetic material (RNA vs. DNA, double stranded vs. single stranded), in their shape (cylindrical, spherical, head-tail), in their mode of entry into the cell, in their biochemistry of replication, in the presence or absence of a lipid envelope, and in other important ways. Even allowing for genetic re-assortment and environmental change, in most cases, it is difficult to visualize how one class of viruses could have given rise to a different one by a process of gradual change. Thus, a transition from one class to another, if it occurred, would seem to have occurred in one sudden jerk, or "saltation,"40 an occurrence explicitly rejected by Darwin and Dawkins (although Dawkins allows the possibility of saltations in rare cases, such as the origin of life).41
The possibilities for gradual evolution
of viral shape are also restricted, but for somewhat more subtle reasons. A
coding triplet of DNA or RNA has a molecular mass of around one thousand
Daltons, while the amino acid it codes for has an average mass of only one
hundred. Since the mass densities of proteins and nucleic acids are roughly the
same, the genetic coding of a given volume of protein requires a ten-fold
greater volume of nucleic acid (RNA or DNA). However, because of the tiny size
of viral particles, and the requirement of complete encapsulation by protein,
viral particles typically contain at least as much protein as nucleic acid.
Thus, the interior volume of the viral protein shell does not suffice to hold
the volume of DNA or RNA required to code for the entire shell! The virus
escapes this "volume paradox" by inducing the host cell to make many
copies of only one or a few small coat proteins which can link together like in
a suit of mail, to completely coat the virus; see Fig. 2.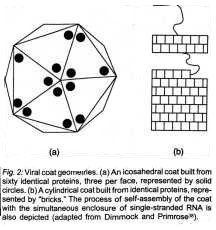
Since individual protein molecules are irregular three-dimensional objects, self-assembly of these into a thermodynamically stable coat imposes certain symmetry requirements on the geometric form the coat can take. Thus, highly symmetric shapes, such as rods, or spherically symmetric "Platonic solids," especially the twenty-sided icosahedron, are preferred. See Fig. 2. A gradual evolutionary transition from one such specialized shape to another thus seems improbable, since intermediate shapes would lack the symmetry of these discrete simple choices, and would therefore fall prey to the "volume paradox." Although recombination events might allow some of the nonshape-coding genetic material from a virus of one shape (e.g., an icosahedron) to be incorporated into the genome of a virus of a different shape (e.g., a rod), it is hard to see how the genetic "modules" that code for shape can themselves change so that one shape is replaced gradually by another different shape. Of course, the protein itself might mutate gradually, but the most stable shape into which the protein self assembles should change discretely and abruptly. Thus, finely-sliced Darwinian evolution of shape is precluded, at least for the smaller viral particles that are constrained by the "volume paradox." This example contradicts Dawkins' claim that small changes in the shape or functioning of an organism are always available to it via small mutations. (In more complex organisms, it is also easy to find examples of changes which cannot be sub-divided into Dawkinsonian "continuous variables." An obvious example is the number of chromosomes, which must be an integral number and differs, e.g., between humans and apes.)
Much more difficult problems attend any transition from one mode of viral replication to another. In one type (Baltimore class IV), the viral genome possesses a single "positive" strand of RNA which must produce its complimentary "negative" strand before messenger RNA is produced to direct protein synthesis. In another type (Baltimore class V), the initial RNA strand is "negative" and can directly induce messenger RNA production. Other RNA viruses (class III) carry double-stranded RNA and reproduce in yet different ways. Finally, RNA retroviruses (class VI) catalyze DNA production via a special enzyme, "reverse transcriptase." Examples of class IV viruses include the common-cold virus, class V include the influenza virus, and class VI the AIDS (HIV) virus. Then there are the single and double-stranded DNA viruses, with yet different reproductive mechanisms.
 Figure
3a illustrates the reproductive
mechanism of lambda-phage, a class I virus which reproduces its double-stranded
DNA directly via a "rolling circle" mechanism. This is entirely
different from the replication mechanism of class IV viruses that use the
infecting single-strand RNA+ as a template to produce RNA-, which then serves as
a template for RNA+ to be packaged in new viruses. See Fig. 3b. The topological
structures of these synthetic pathways are inherently distinct, and the steps in
each pathway are few. Hence, a change in the pathway would need to take place by
discreet additions or subtractions of pathway steps. And, since the product of
one step is the RNA, DNA, or protein needed for the next step in the pathway,
the disappearance or replacement of one step would certainly have to be
accompanied by other simultaneous changes to maintain continuity of the
production cycle. Thus, while the rate at which a single step in the pathway is
carried out might vary as a result of small mutations, complete removal or
insertion of a step in the pathway is an intrinsically large evolutionary step
that cannot be broken down into a series of smaller ones. The Darwinian
requirement of "gradual, continuous improvement" (or descent with
modification) would not seem to be available.
Figure
3a illustrates the reproductive
mechanism of lambda-phage, a class I virus which reproduces its double-stranded
DNA directly via a "rolling circle" mechanism. This is entirely
different from the replication mechanism of class IV viruses that use the
infecting single-strand RNA+ as a template to produce RNA-, which then serves as
a template for RNA+ to be packaged in new viruses. See Fig. 3b. The topological
structures of these synthetic pathways are inherently distinct, and the steps in
each pathway are few. Hence, a change in the pathway would need to take place by
discreet additions or subtractions of pathway steps. And, since the product of
one step is the RNA, DNA, or protein needed for the next step in the pathway,
the disappearance or replacement of one step would certainly have to be
accompanied by other simultaneous changes to maintain continuity of the
production cycle. Thus, while the rate at which a single step in the pathway is
carried out might vary as a result of small mutations, complete removal or
insertion of a step in the pathway is an intrinsically large evolutionary step
that cannot be broken down into a series of smaller ones. The Darwinian
requirement of "gradual, continuous improvement" (or descent with
modification) would not seem to be available.
Therefore, each viral type (selected, e.g., from Baltimore classes I through VI) has a fundamentally different reproduction strategy. A gradual shift from one viral type to another by way of accumulated point mutations, or even re-assortments, is hard to imagine. Rather, the collection of all possible viral forms appears to be a set of discrete points in configurational space and not a continuum of possibilities. If so, evolution from one state to the next would have to occur by "saltation," that is, large hops, rather than by the slow, gradual transitions envisioned by Darwin and Dawkins.
Recognizing these difficulties, virologists have speculated that some viral types originated completely independently of each other. In one scenario, viruses are genetic components that escaped from a cell.44 In this view, each different viral type is the offspring of the cell type that it now infects. Another scenario holds that viruses are the end result of the degeneration of free-living cells. One might also speculate that viruses arose during the "primordial" period of molecular evolution, which is presumed to have preceded the emergence of cellular life. In this third scenario, as biochemical networks became organized into enclosed communities or cells, the portions of the network rejected by the cells became "outlaws," surviving by attacking and ransacking the cells from which they were excluded. One might also speculate that viral particles originated from cancerous cells. Cells become cancerous when their regulatory control breaks down, leading to runaway reaction networks. A self-sustaining reaction network compact enough to become independent of the cell from which it originated might become a virus, if the network is able also to produce a suitable protective protein coat.
While such scenarios make for interesting speculation, they stand outside the boundaries of hard, empirical science. For example, while it is true that some viruses, such as hepadna viruses, can produce cancer,45 this does not mean that the reverse has ever occurred. Furthermore, whichever scenario one might favor, each constitutes non-Darwinian evolution; that is, evolution not by small changes as Darwin envisioned, but by "saltation," that is, large changes whose occurrence is far from commonplace, very unlike the small mutations upon which Darwinism rests. Thus, even in the most favorable of circumstances of the high-speed mutations and short reproductive cycle of RNA viruses, scientific demonstration of the evolutionary radiation of different viral strains from a single progenitor seems to be beyond the reach of known empirical science. Appearance of new viral forms seems to depend on highly fortuitous circumstances, odd twists of nature, and freakish events that are both unpredictable and impossible to reconstruct with any confidence.
The Tree or the Briar Patch?
Nor do these conclusions apply to viruses only. Recent genomic
sequencing shows that the simplest bacterial life forms that are considered to
be near the base of the tree of life have genomes that are incompatible with the
simple "descent-with-modification" formula of Darwin and Dawkins. In
Darwinism, family relationships among organisms must be that of a genealogical
tree--that is, a branching structure. This point is emphasized forcefully by
Dawkins:
It follows from the idea of evolution that there is one uniquely correct branching family tree of all living things, ... [which] has the property of perfect nesting. We write the names of any set of animals on a large sheet of paper and draw rings round related sets ... Never, not on a single solitary occasion, will the rings that we draw intersect each other.46
Yet, near the base of the tree of life, it is now known that the rings do intersect each other.
Life at its most basic is currently divided into three kingdoms: eukaryotes, simple bacteria, and "archaea." (Archaea are known for living in extreme environments, such as boiling-hot, deep-sea thermal vents.) The relationships among single-celled organisms in these three different kingdoms of life must be graphed as a network, rather than a tree. Thus, e.g., eukaryotic cells have large numbers of informational genes that are closely related to archaea, while the operational genes are more closely related to those of bacteria.47 Since bacteria and archaea are traditionally regarded as more ancient than the more advanced eukaryotic cells, the presence of some genes in eukaryotes that are unique to bacteria, and other genes that are unique to archaea, could only mean that the eukaryotes are "chimeric"--that is, a fusion of two earlier life forms! Extensive study of the genomes of these simple organisms are thus not consistent with Darwinian descent, but instead can only be explained by postulating the repeated transfer of whole genes and even large clusters of genes from organism to organism. To account for the genome of the bacterium E. Coli, e.g., one must postulate that no fewer than 755 of its genes have been transferred to it in at least 230 "lateral" transfers from other organisms.48 Thus, "gene swapping" may have "turned the tree of life into a tangled briar whose lineages may be next to impossible to discern," says Pennisi in a 1999 Science article entitled "Is it time to uproot the tree of life?"49
One might be tempted to relegate such non-Darwinian processes to the simplest of life forms, such as viruses, bacteria, archaea, and the like. However, the explosion in genetic information is also turning up similar examples in higher organisms. Dawkins himself notes one example. The gene for hemoglobin (a key protein in the red blood cells of higher animals) has been found to exist in plants of the pea family, and hemoglobin itself exists in the roots of this plant! Hemoglobin has not been found in any other plant families, and so could not have been passed down to this species by "descent with modification." Instead, it must have been transferred to the pea plant directly from an animal, perhaps by a viral carrier. With the rapid sequencing of genomes, many more genes are turning up in "the wrong place." For example, the bacterium, Deinococcus radiodurans, contains several genes found up to now only in plants. Another bacterium, mycobacterium tuberculosis, contains at least eight human genes.50
Furthermore, there are many examples of "convergent evolution," in which animals in remote families of life are uncannily similar to each other in many respects. An example is the Australian marsupial "mole," which, except for its pouch, is almost indistinguishable from an ordinary placental mole. Other examples include a marsupial "saber-toothed tiger" and a marsupial "anteater," each very similar to its evolutionary, remote placental counterpart. As long as these similarities are purely external, one can explain them (as Dawkins does) as the result of a similar selection pressure acting on initially very different organisms, leading them to acquire similar traits.51
One such trait is the eye, which Dawkins believes evolved on at least forty separate occasions.
Recent
genomic sequencing shows that the simplest
bacterial life forms that are considered to be near
the base of the tree of life have genomes that are incompatible with the simple
"descent-with-modification" formula of Darwin and Dawkins.
Much harder to explain, however, are extreme similarities in the biochemical machinery adopted by diverse organisms as they shape themselves to their environment. In the case of the eye, e.g., it has now been found that the gene that triggers its formation in the embryos of animals as diverse as insects and mice is almost identical! This gene is called "eyeless" because its deletion from the genome of an organism results in failure of that organism to produce eyes during embryonic development.52 However, the presumed Darwinian structure of the "tree of life," and the absence of eyes in some of the species that are considered evolutionary links between species with eyes, implies that eye formation in these separate organisms occurred independently! Thus, the "random" mutations that led on multiple separate occasions to eye formation appear to have repeatedly recruited the same gene "eyeless" to trigger eye formation. What would have happened had this particular gene never been invented by evolution? Darwin and Dawkins say that nature is resourceful; some other gene could equally well have been mutated to form eyes. But the evidence strongly suggests that paths to the eye that do not involve the "eyeless" gene are rare or nonexistent; otherwise nature would not have gone back repeatedly to the same gene to initiate eye formation.
A similar story has recently emerged regarding the evolution of "cryptochromes," which are photoreceptor proteins in plants and animals.53 As in the case of "eyeless," the same evolutionary pathway seems to have been followed repeatedly, even at the biomolecular level, contradicting Dawkins' statement that "it is vanishingly improbable that exactly the same evolutionary pathway should ever be travelled twice."54 The specificity of the evolutionary steps required for these transformations calls into question Dawkins' assertion that the mutations necessary for evolution were "bound to be forthcoming." Rather, instead of evolving gradually by trial-and-error "invention," organisms seem to have resorted to "espionage," stealing heavily from each other across vast distances in the family "tree."
The analogy of monkeys with typewriters, therefore, is not an apt metaphor for evolution. Rather, one must suppose that the monkeys are stealing whole pages from each other's typewriters. In such a version of evolution, Lincoln's Gettysburg address would be produced not by an accumulation of small typing errors, but by random splicing together of the Magna Carta and Beowulf.
Philosophies
Of course, this is just what many believe happened. That is,
"mix-and-match" transfer of large chunks of genetic material from one
organism to another is increasingly thought of as a vital part of the
evolutionary process. It remains to be seen how much of evolution must be
accounted for by non- Darwinian "borrowing," as more and more genomes
are sequenced. Of course, "non-Darwinian" does not necessarily imply
"improbable," let alone "miraculous." Broadly speaking, one
can look on the question of origins in one of five ways, allowing for gradations
within and between each way.
1. Darwinian Evolution: Accumulation of small mutations by natural selection. The pace of evolution may vary from "slow" (little change over millions of years) to "fast" (substantial change over millions of years), as long as no large step occurs in a single generation.
2. Random Saltation: Major changes by large "jumps" in a single generation, either by "borrowing" genetic material from another organism, frame shifts in the genome, increase in chromosome number, etc. While these changes are very rarely favorable, enough time is available to make them probable.
3. "Anthropic" Saltation: As in position 2, except that the changes are so rarely favorable that large-scale evolution is unlikely to occur in any one universe. With infinite numbers of possible universes, any universe with observers (like ours) is inevitably one in which these overwhelmingly unlikely events have occurred.
4. "Providential" Saltation: As in position 3, except that the extremely improbable events in a single universe were foreordained and planned by God, who "engineered" them without violating physical laws, either through setting of initial conditions, exploiting of quantum uncertainty, or other, perhaps unknowable, methods.
5. Miraculous Creation: Transformation or creation of organisms by divine processes that supercede physical laws or principles.
Of course, "hybrid" positions combining two or more of the above are possible. Dawkins's position is 1, with some admixture of 2 to account for the origin of life. Philosophically, positions 1 through 3 are compatible with atheism, while positions 4 and 5 obviously are not. However, empirically, position 3 cannot be distinguished from position 4. Both involve events that are probabilistically impossible within a single universe. Practically, it may well be impossible (at least in the short run) to distinguish position 2 from position 3, since both involve events whose probability seems to be impossible to estimate. Likewise, "moderate" forms of position 5 may not really be distinguishable empirically from position 4, although certainly a radical creation event, such as a literal "six-day" creation, would be distinguishable from position 4. Thus, from Dawkins' point of view, one of the main appeals of evolutionary theory, namely that it renders God redundant, would be put at risk were even position 2 to be considered more likely than his own position 1. This may account for his strong attachment to the strict Darwinian position. It must be noted, however, that he honestly owns up to some limitations of this position when forced to. Therefore, it will be most interesting to watch the "evolution" of his position and that of other Darwinists, as the floodgates of genomics open.
Summary
The backside of Mount Improbable is not the "gently sloping meadow" of
Dawkins' imagination. Instead, consideration of even the simplest self-
replicating entities, RNA viruses, shows that even the backside is a rugged
landscape filled with wide canyons and harrowing ledges. The climb up this
mountain requires not merely steady walking, but fortuitous landslides to fill
in the canyons, and timely earthquakes to split the ledges. Since the earth is
some 4.5 billion years old, such "coincidences" cannot logically be
ruled out. However, the elevation of evolutionary theories of origins from the
speculative or "folk-lore" status they held before Darwin to the
unquestioned acceptance that they have since been accorded is now being eroded
by the advances in genomics and biochemistry. We should no longer place
Darwinism as a universal theory of the origin of species alongside empirically
established scientific principles such as the laws of thermodynamics.
Notes
1Richard Dawkins, The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe Without Design (New York: W.W. Norton & Co, 1987).
2------, Climbing Mount Improbable (New York: W. W. Norton & Co, 1996).
3Ibid., 73.
4Ibid., 147.
5Charles Darwin, The Origin of Species, sixth ed. (Amherst, NY: Prometheus Books, 1991), 139.
6Dawkins, Climbing Mount Improbable, 91.
7------, The Blind Watchmaker, 77.
8Ibid., 78.
9Ibid.
10Ibid., 79.
11Ibid.
12Ibid.
13Ibid., 83-4.
14Ibid., 85-6.
15------, Climbing Mount Improbable, 138-97.
16D.-E. Nilsson and S. Pelger, "A Pessimistic Estimate of the Time Required for an Eye to Evolve," Proceedings of the Royal Society, London, B 256 (1994): 53-8.
17Dawkins, The Blind Watchmaker, 47-8.
18------, Climbing Mount Improbable, 75.
19L. Stryer, Biochemistry, 3rd ed. (New York: W. H. Freeman and Company, 1988), 107.
20J. W. Drake, "Rates of Spontaneous Mutations Among RNA Viruses," Proceedings of the National Academy of Science 90 (1993): 4171-5; and R. A. Beckman and L. A. Loeb, "Multi-stage proofreading in DNA replication," Quarterly Reviews of Biophysics 26 (1993): 255-331.
21E. Strauss, "Can Mitochondrial Clocks Keep Time?" Science 283 (1999): 1435-8.)
22Dawkins, The Blind Watchmaker, 57.
23E. Domingo and J. J. Holland, "RNA Virus Mutations and Fitness for Survival," Annual Reviews of Microbiology 51 (1997): 151-78; D. S. Steinhauer and J. J. Holland, "Rapid Evolution of RNA Viruses," Annual Reviews of Microbiology 41 (1987): 409-33; J. W. Drake, "Rates of Spontaneous Mutations Among RNA Viruses," Proceedings of the National Academy of Science 90 (1993): 4171-75; P. Palese, "Evolution of Influenza and RNA Viruses," in Emerging Viruses, ed. S. S. Morse (New York: Oxford University Press, 1993); E. Domingo, C. EscarmÌs, N. Sevilla, A. Moya, S. F. Elena, J. Quer, I. S. Novella, and J. J. Holland, "Basic Concepts in RNA Virus Evolution," The FASEB Journal 10 (1996): 859-64.
24N. J. Dimmock and S. B. Primrose, Modern Virology, 4th ed. (Oxford, UK: Blackwell Science, 1994), 337.
25Domingo and Holland, "RNA Virus Mutations and Fitness for Survival," 153.
26I. S. Novella, J. Quer, E. Domingo, and J. J. Holland, "Exponential Fitness Gains of RNA Virus Populations Are Limited by Bottleneck Effects," Journal of Virology 73 (1999): 1668-71.
27Ibid.
28Ibid.
29E. A. Duarte, D. K. Clarke, A. Moya, E. Domingo, and J. J. Holland, "Rapid Fitness Losses in Mammalian RNA Virus Clones Due to Muller's Ratchet," Proceedings of the National Academy of Sciences, 89 (1992): 6015-19.
30E. A. Duarte, I. S. Novella, S. Ledesma, D. K. Clarke, A. Moya, S. F. Elena, E. Domingo, and J. J. Holland, "Subclonal Components of Consensus Fitness in an RNA Virus Clone," Journal of Virology 68 (1994): 4295-301; and Domingo, et al., "Basic Concepts in RNA Virus Evolution," 859-64.
31H. J. Muller, "The Relation of Recombination to Mutational Advance," Mutation Research 1 (1964): 2-9; and Domingo, et al., "Basic Concepts in RNA Virus Evolution," 859-64.
32Domingo and Holland, "RNA Virus Mutations and Fitness for Survival," 165; and M. A. MartÌnez, J. Dopazo, J. Hernadez, M. G. Mateu, and F. Sobrino, "Evolution of the Capsid Protein Genes of Foot-and-Mouth Disease Virus: Antigenic Variation Without Accumulation of Amino Acid Substitutions over Six Decades," Journal of Virology 66 (1992): 3557-65.
33Domingo and Holland, "RNA Virus Mutations and Fitness for Survival," 151-78.
34S. S. Morse, "Examining the Origins of Emerging Viruses," in Emerging Viruses, ed. S. S. Morse (New York: Oxford University Press, 1993), 10-28.
35Ibid., 15.
36Dimmock and Primrose, Modern Virology, 286.
37C. S. Hahn, S. Lustig, E. G. Strauss, and J. H. Strauss, "Western Equine Encephalitis Virus Is a Recombinant Virus," Proceedings of the National Academy of Sciences 85 (1988): 5997-6001.
38J. H. Strauss, "Recombination in the Evolution of RNA Viruses," in Emerging Viruses, ed. S. S. Morse (New York: Oxford University Press, 1993), 241-51.
39J. P. Icenogle, P. Sathya, D. I. Miller, R. A. Tucker, and W. E. Rawls, "Nucleotide and Amino Acid Sequence Variation in the L1 and E7 Open Reading Frames of Human papillomavirus Type 6 and Type 16," Virology 184 (1991): 101-7; and T. C. Yamada, C. M. Wheeler, A. L. Halpern, A-C. M. Stewart, A. Hildesheim, S. A. Jenison, "Human papillomavirus Type 16 Variant Lineages in United States Populations Characterized by Nucleotide Sequence Analysis of the E6, L2, and L1 Coding Segments," Journal of Virology 69 (1995): 7743-53.
40A large step, or jump, in a single generation is known as a "saltation." Evolution by saltations is firmly rejected by both Darwin and Dawkins (with some exceptions, in the case of Dawkins). Now that we know the genetic basis of life, we can discern two kinds of saltation. One of these was recognized, and dismissed, by Darwin. It is a large jump in "phenotype"--that is, the outward form and function of the organism. These are considered to be virtually impossible, because any large jump, if it is random, is almost certain to be a disaster for the organism. Hitting a malfunctioning watch with a sledge hammer may produce change, but not likely for the better. Such "phenotype" saltations need not be produced by a large change in the underlying genetic code. For instance, replacement of a single amino acid by another can sometimes cause the resulting protein to fold improperly, or to fail to function, with sometimes disastrous consequences. Most cases of the disease cystic fibrosis are produced by a deletion of the three-base codon for the single amino acid phenylalanine in a protein that controls the regulation of salt concentration in cells. The external change produced by this mutation is a large one, and is highly unfavorable to the survival of the person possessing it. Another example of a small change in the DNA that produces large phenotypic changes is a "frame shift." This occurs when one base pair of the DNA sequence is either accidentally left out or added to a DNA sequence. Since the sequence is read in triplets, such a deletion or insertion will result in a shift to the left or to the right by one letter in the assignment of codons. Hence, downstream of the "error," practically every amino acid assignment will be altered, with massive changes to the protein, and likely catastrophic consequences for the organism. A second kind of saltation is a "genotypic" saltation, wherein large changes randomly occur within protein-coding regions of the genome. Such a saltation is unlikely to produce favorable change because, if it is truly random, it will almost certainly produce a large phenotypic change, and therefore be unfavorable. In principle, a "genotypic" saltation could produce only a small change in "phenotype," if the genotypic changes somehow cancel each other out, leading to only a small change in phenotype. But such a near-perfect cancellation is unlikely to arise from a random change, just as it is unlikely that a large, random, phenotypic change will be beneficial.
41The possibility of impediments to gradualism are acknowledged by Dawkins. For example, despite the supposed ease and rapidity of evolution of the camera eye from a simple photoreceptor cell, the inefficient compound eye of insects has been unable, after millions of years, to evolve into the camera eye. Dawkins suggests that the inverted image of the camera eye represents an inherently discontinuous change from the right-side-up image of the more primitive compound eye. "Finding an intermediate between these two is a tough proposition, to put it mildly," he writes. Thus, the compound eye, though non-optimal, might represent a foothill, separated by an impassable valley from the true optimal eye, which resides on the peak of Mount Improbable itself. The obvious question, then, is "How common are such impasses?" or "How smooth really is the backside of Mount Improbable?"
42Dimmock and Primrose, Modern Virology, 44.).
43Ibid., 106, 118.
44Ibid., 335-7.
45Ibid., 256-75.
46Dawkins, The Blind Watchmaker, 239.
47E. Pennisi, "Is it Time to Uproot the Tree of Life?" Science 284 (1999): 1305-7.
48J. A. Lake, R. Jain, and M. C. Rivera, "Mix and Match in the Tree of Life," Science 283 (1999): 2027-8.
49Pennisi, "Is it Time to Uproot the Tree of Life?" 1305-7.
50Ibid.
51Dawkins, The Blind Watchmaker, 105.
52C. S. Zuker, "On the Evolution of Eyes: Would You Like It Simple or Compound?" Science 265 (1994): 742-3.
53A. R. Cashmore, J. A. Jarillo, Y.-J. Wu, and D. Liu, "Cryptochromes: Blue Light Receptors for Plants and Animals," Science 284 (1999): 760-5.
54Dawkins, The Blind Watchmaker, 94.