
Keith B. Miller
Department of Geology
Kansas State University, Manhattan, KS 66506
The recognition and interpretation of patterns in the fossil record require an awareness of the limitations of that record. Only a very small fraction of the species that have lived during past geologic history is preserved in the rock record. Most marine species are soft-bodied, or have thin organic cuticles, and are essentially unpreservable except under the most extraordinary conditions. Furthermore, the destructive processes active in most marine environments prevent the preservation of even shelled organisms under normal conditions. Preservational opportunities are even more limited in the terrestrial environment. Most fossil vertebrate species are represented by no more than a few fragmentary remains. Because of the preservational biases of the fossil record, paleontologists must reconstruct evolutionary relationships from isolated branches of an originally very bushy tree.
The process of describing and classifying organisms introduces its own patterns into the taxonomic hierarchy. First, because organisms must be placed in one group or another, taxonomy gives the impression of discontinuity. Secondly, the placement of species into higher taxa is done retrospectively; that is, by looking backward through time. The evolutionary significance of particular morphologic transitions is only recognized because of the subsequent success of particular lineages. The defining characters of higher taxa are thus a consequence of history, and do not represent some objective scale of the magnitude of morphologic divergence. Closely-related species from two different higher taxa may actually be more similar in morphology than two distantly-related species belonging to the same group.
Because new character states are added over geologic time, the morphology of species within a higher taxonomic group becomes less divergent toward the point of origin of that group. In addition, species appearing early in the history of a taxon approach more closely the morphology of species from other closely related higher taxa, often to the extent that their taxonomic assignment is uncertain. Transitional forms between higher taxa are thus a common feature of the fossil record, although continuous fossil lineages are rarely if ever preserved. Evidence from the fossil record is consistent with a wide range of proposed evolutionary mechanisms.
The fossil record provides persuasive evidence for macroevolutionary change and common descent. The pattern of appearance of fossil species through geologic time is critical for reconstructing evolutionary relationships. In addition, the fossil record may also contribute to our understanding of the tempo and mode of evolution, and help select between competing macroevolutionary theories.
However, before the fossil record can be applied to these questions, two critically important topics need to be addressed. The first concerns the completeness and resolution of the fossil record, and the second concerns taxonomic procedures. Taxonomy refers to the methods by which species are defined and grouped into a hierarchy of categories.
There are two opposite errors which need to be countered about the fossil record: (1) that it is so incomplete as to be of no value in interpreting patterns and trends in the history of life, and (2) that it is so good that we should expect a relatively complete record of the details of evolutionary transitions within most lineages.
What then is the nature of the fossil record? It can be confidently stated that only a very small fraction of the species that once lived on Earth has been preserved in the rock record and subsequently discovered and described by science. Our knowledge of the history of life can be put into perspective by a comparison with our knowledge of living organisms. About 1.5 million living species have been described by biologists, while paleontologists have catalogued only about 250,000 fossil species representing over 540 million years of Earth history (Erwin, 1993)! Why such a poor record?
Soft-bodied or thin-shelled organisms have little or no chance of preservation, and the majority of species in living marine communities are soft-bodied. Consider that there are living today about 14 phyla of worms comprising nearly half of all animal phyla, yet only one, the Annelida, has a significant fossil record. The inadequacy of the fossil record to preserve with any completeness the evolutionary history of soft-bodied organisms can be illustrated by the Conodonta. Originally assigned to their own phylum, they are now believed to belong to the cordates. These soft-bodied animals are represented by tiny tooth-like phosphatic fossils which are very abundant in sedimentary rocks extending over about 300 million years of Earth history, and have a worldwide distribution. Conodonts are a very important group of marine fossils for paleontologists, yet until only very recently the organism to which they belonged was completely unknown. Specimens of the worm-like conodont animal have now been discovered in Carboniferous, Ordovician, and Silurian rocks (Briggs et al., 1983; Mikulic et al., 1985; Aldridge & Purnell, 1996). Only a handful of specimens is now known from a very large and diverse group of marine animals known to be extremely abundant and widespread over a tremendous length of time!
The discovery of new soft-bodied fossil localities is always met with great enthusiasm. These localities typically turn up new species with unusual morphologies, and new higher taxa are built from a few specimens! Such localities are also erratically and widely spaced in geologic time between which essentially no soft-bodied fossil record exists.
Even those organisms with preservable hard parts are unlikely to be preserved under "normal" conditions. Recent studies of the fate of clam shells in shallow coastal waters reveal that shells are rapidly destroyed by scavenging, boring, chemical dissolution, and breakage. Rare events such as major storms appear to be required to incorporate shells into the sedimentary record. Getting terrestrial vertebrate material into the fossil record is even more difficult.
The limitations of the vertebrate fossil record can be easily illustrated. The famous fossil Archaeopteryx, occurring in a rock unit renowned for its fossil preservation, is represented by only seven known specimens, of which only two are essentially complete. Considering how many individuals of this genus probably lived and died over the thousands or millions of years of its existence, these few known specimens give some feeling for how few individuals are actually preserved as fossils and subsequently discovered. Yet this example actually represents an unusual wealth of material. The great majority of fossil vertebrate species are represented by only very fragmentary remains, and many are described on the basis of single specimens or from single localities. Complete skeletons are exceptionally rare. For many fossil taxa, particularly small mammals, the only fossils are teeth and jaw fragments. If so many fossil vertebrate species are represented by single specimens, the number of completely unknown species must be enormous!
In addition to these preservational biases, the erosion, deformation, and metamorphism of originally fossiliferous sedimentary rocks have eliminated significant portions of the fossil record over geologic time. Furthermore, much of the fossil-bearing sedimentary record is hidden in the subsurface, or located in poorly accessible or little studied geographic areas. For these reasons, of those once living species actually preserved in the fossil record, only a small portion has been discovered and described by science.
Because of the biases of the fossil record, the most abundant and geographically widespread species of hard part-bearing organisms would tend to be best represented. Also, because evolutionary change is probably most rapid within small isolated populations, species within rapidly evolving lineages are less likely to be preserved in the fossil record. In addition, the completeness of the fossil record improves up the taxonomic hierarchy (Erwin, 1993). A smaller proportion of once-living species is preserved than genera, of genera than families, of families than orders, etc. As a result we can better discern the general patterns of evolutionary change than the population-by-population or species-by-species transitions.
Given the limitations and biases discussed above, what should be expected from the fossil record? The situation is not as bleak as it may appear from my previous comments. Exceptional deposits, such as the Burgess Shale, Solnhofen Limestone, and Green River Shale, do provide surprisingly detailed glimpses of once living communities. These rare cases of exceptional preservation (fossil lagerstätten) are essentially snapshots in the history of life and are invaluable in gaining a more comprehensive picture of ancient communities. They also provide some of the most detailed anatomical data.
More commonly, thick sequences of fossiliferous rocks can enable selected skeleton or shell-bearing taxa to be examined at closely-spaced intervals. These localities provide opportunities to study patterns of evolutionary change within isolated lineages. Important information can be gained on morphologic change within species populations, and transitions between species and, rarely, even genera can be examined (Fig. 1). However, the time interval recorded by continuous series of closely-spaced fossil populations is limited because of changing environmental, depositional, and preservational conditions.
Figure 1. Changes in the shape of molar teeth of the Early Eocene mammal Hypsodus, showing evolutionary transitions from species to species within a genus. (From Gingerich [1976], reprinted with permission of the American Journal of Science.)
Speciation events appear to take place primarily in small isolated peripheral populations. Therefore to catch a population "in the act" requires the fortuitous sampling of the particular geographic locality where the changes occurred. Even within well-preserved fossil series it is usually difficult to distinguish the record of speciation occurring within a particular depositional basin (or environment) from the effects of immigration of new species from outside that basin. For this and other reasons, well-documented and widely-accepted examples of speciation in the fossil record are few (for an example, see Gingerich, 1976).
The expectation, therefore, is for the preservation of isolated branches on an originally very bushy, evolutionary tree. A few of these branches (lineages) would be fairly complete, while most are reconstructed with only very fragmentary evidence (Fig. 2). While the details are missing, a general understanding of the large-scale patterns and trends in evolutionary history should be discernible. Evolutionary trends over longer periods of time and across greater morphologic transitions can be followed by reconstructing morphological sequences. Morphological transitions can be recognized in the fossil record that cross all levels of the taxonomic hierarchy.

Figure 2. The effects of an incomplete fossil record on the reconstruction of evolutionary relationships. (A) This branching tree (phylogeny) represents the actual pattern of evolutionary relationships. (B) The actual preserved record of species in the fossil record might look something like this. (C) This branching tree represents a possible reconstruction of the evolutionary tree based on the fossil evidence. Note that the general pattern of relationships is preserved, but that errors have been made with regard to specific ancestor-descendant relationships.
Taxonomy, the process of classifying living and fossil organisms, produces its own patterns which order the diversity of life. It is thus important to recognize that names do much more than describe nature: they also interpret it. There is considerable ferment now within the field of taxonomy because of conflicting philosophies of classification, and different perceptions of which patterns in the history of life should be reflected in the taxonomic hierarchy (Eldredge & Cracraft, 1980; Schoch, 1986). Higher taxa can be either artificial groupings of species with similar morphologies (evolutionary grades), or "natural" groups sharing derived characteristics inherited from a common ancestor (monophyletic taxa or clades).
The Linnean classification system is hierarchical, with species grouped into genera, genera into families, families into orders, etc. This system reflects the discontinuity and hierarchy observed among living organisms. However, "this system leads to the impression that species in different categories differ from one another in proportion to differences in taxonomic rank" (Carroll, 1988, p. 578). This impression is false. Higher taxa are distinct and easily recognizable groups only when we ignore the time dimension of the history of life. When the fossil record is included, the boundaries between higher taxa become blurred during the major morphological radiations associated with the appearance of new higher taxa. Even in the modern world, discontinuity is not as great as it may appear superficially. In practice, species are often not easily recognized, and accepted species definitions cannot always be applied.
Another common misperception is that the origin of higher taxa does not take place at the level of populations and species. If the concept of common descent is accepted, then transitions between higher level taxonomic categories must also be species transitions (Fig. 3). This is recognized by all evolutionary paleobiologists, even those who stress the significance of the origin of phyla and classes (Valentine, 1992). Therefore, the more complete the fossil record of the origin and early radiation of higher taxa the more similar the transitional species, and the more difficult it is to determine their taxonomic assignments. Species placed into two different higher taxa may thus have very similar morphologies.
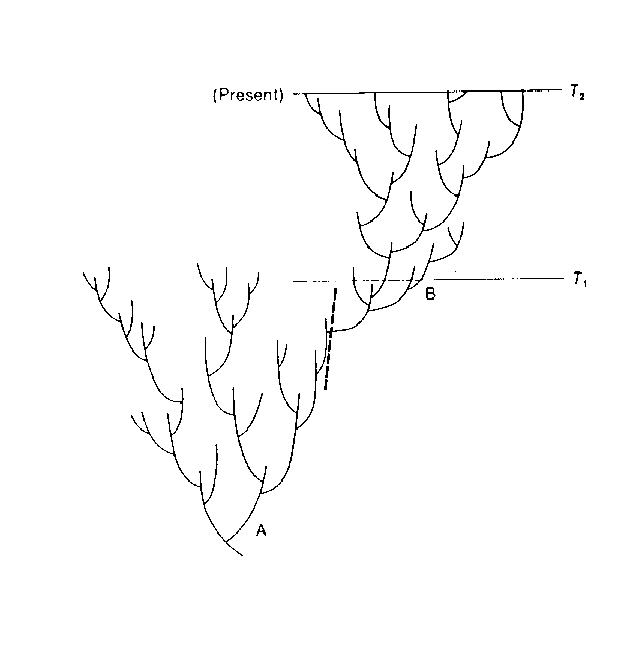
Figure 3. Pattern of phylogeny in which one clade (or higher taxon) emerges from another. In retrospect (time T2), the two clades are seen as being distinct, and the phylogeny is divided at the position of the heavy, dashed bar into taxa A and B. A taxonomist living at time T1, however, would have recognized only a single clade and would have grouped the entire phylogeny that had developed by that time into a single taxon (A). (From Macroevolution: Pattern and Process by Stanley © 1979 by W.H. Freeman and Company, used with permission. All rights reserved.)
The character states used to define higher taxa are determined retrospectively. That is, they are chosen based on a knowledge of the subsequent history of the lineages possessing those traits. They do not reflect the attainment of some objective higher level of morphologic innovation at the time of their appearance. Also, all the features subsequently identified with a particular higher taxon do not appear in a coordinated and simultaneous manner but as character mosaics within numerous closely-related species lineages, many of which are not included in the new higher taxon. In addition, as discussed above, the species associated with the origin and initial radiation of a new taxon are usually not very divergent in morphology. Were it not for the subsequent evolutionary history of the lineages, species spanning the transitions between families, orders, classes, and phyla would be placed in the same lower taxon (Fig. 3).
Based on the above discussion, a transitional form is simply a fossil species that possesses a morphology intermediate between that of two others belonging to different higher taxa. Such transitional forms commonly possess a mixture of traits considered characteristic of these different higher taxa. They may also possess particular characters that are themselves in an intermediate state. During the time of origin of a new higher taxon, there are often many described species with transitional morphologies representing many independent lineages. It is usually very difficult if not impossible to determine which, if any, of the known transitional forms actually lay on the lineage directly ancestral to the new taxon. For this reason, taxonomists commonly have difficulty defining higher taxa, and assigning transitional fossil species to one or the other taxon. But, although the details may elude us, the patterns of evolutionary change are in many cases well recorded in the fossil record.
As stated above, the diversity of life appears much more discontinuous when viewed at any given point in time, than it does when viewed through time. For a given time slice through the tree of life, transitions between taxa are seen only where the slice intersects the branching points of lineages. Once a lineage is split, its branches continue to evolve and diverge such that their morphological (and genetic) distance increases and they become more readily distinguished taxonomic entities. When looking backward through time using the fossil record, it is found that representatives of different higher level taxa become more "primitive," that is have fewer derived characters, and appear more like the primitive members of other closely related taxa. As a result, for lineages with a good fossil record, the appearance of a new higher taxon is associated with the occurrence of species whose taxonomic identities are uncertain or whose morphologies converge closely on that of the new higher taxon. Such patterns are found repeatedly by paleontologists.
A longstanding misperception of the fossil record of evolution is that fossil species form single lines of descent with unidirectional trends. Such a simple linear view of evolution is called orthogenesis, and has been rejected by paleontologists as a model of evolutionary change (MacFadden, 1992). The reality is much more complex than that, with numerous branching lines of descent and multiple morphologic trends (Fig. 4). The fossil record reveals that the history of life can be understood as a densely branching bush with many short branches (short-lived lineages). The well-known fossil horse series, for example, does not represent a single continuous evolving lineage (MacFadden, 1992). Rather it records more or less isolated parts of an adapting and diversifying limb of the tree of life. While incomplete, this record provides important insights into the patterns of morphological divergence and the modes of evolutionary change.

Figure 4. Comparison of a single direct line of descent (orthogenesis) with a branching phylogeny. Diversification is such an important feature of the history of life that orthogenesis is probably very rare. Fossils from a chronological series thus do not represent direct ancestor-descendant relationships, but individual branches. (From MacFadden [1992], reprinted with permission of Cambridge University Press).
Interestingly, some critics of evolution view the record of fossil horses from "Eohippus" (Hyracotherium) to Equus as trivial (Denton, 1985). However, that is only because the intermediate forms are known (Fig. 5, 6). Without them, the morphologic distance would appear great. "Eohippus" was a very small (some species only 18 inches long) and generalized herbivore (probably a browser). Besides the well-known difference in toe number (four toes at front, three at back), "Eohippus" had a narrow elongate skull with a relatively small brain and eyes forward in the skull. It possessed small canine teeth, premolars, and low-crowned simple molars. Over geologic time and within several lineages, the skull became much deeper, the eyes moved back, and the brain became larger. The incisors were widened, premolars were altered to molars, and the molars became very high-crowned with a highly complex folding of the enamel (Evander, 1989; McFadden, 1988).
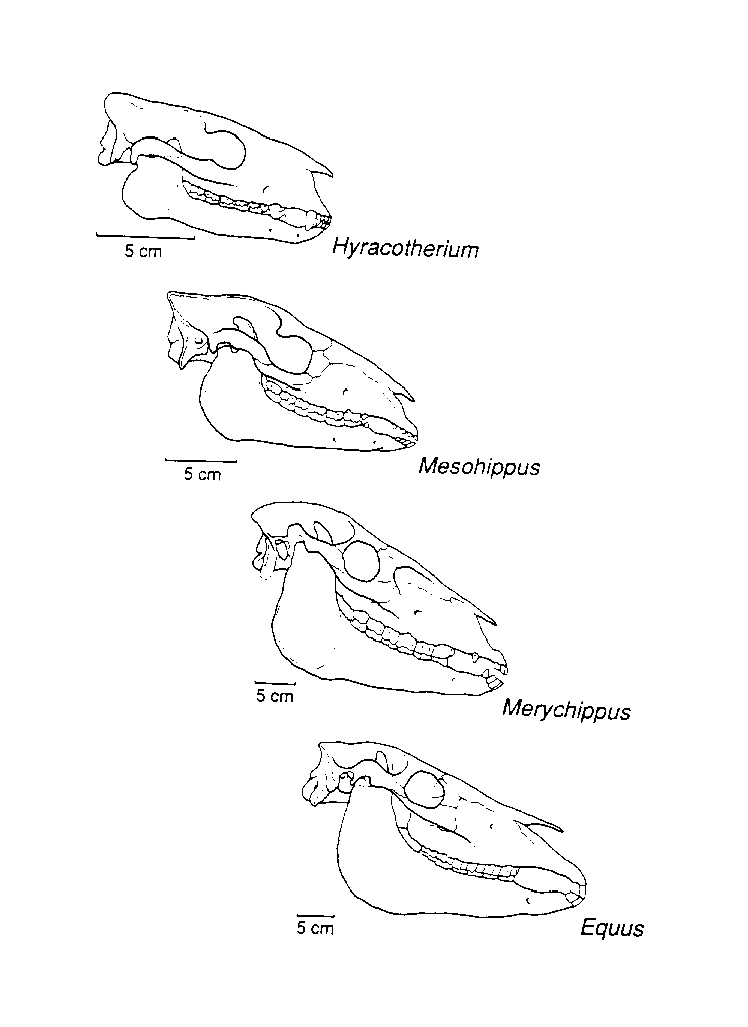
Figure 5. Fossil horse series from Hyracotherium ("Eohippus") to Equus showing changes in skull proportions associated with an adaptive shift from browsing to grazing. This sequence shows a chronological sequence of genera within the perissodactyl family Equidae from the Eocene to the Recent. (From MacFadden [1992], reprinted with permission of Cambridge University Press).
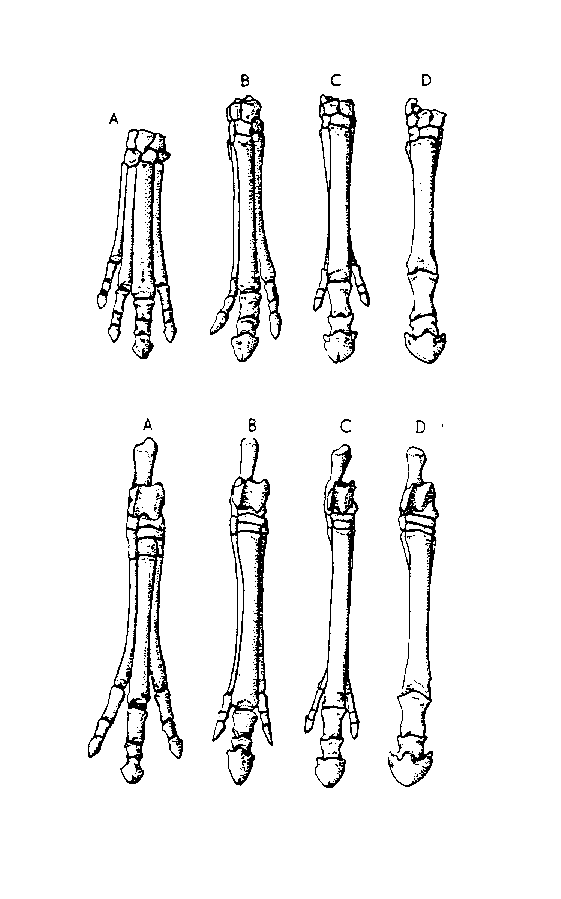
Figure 6. Stages in horse evolution showing the reduction in the number of toes and foot bones. Forefeet above, hind feet below. (A) Hyracotherium, a primitive early Eocene horse with four toes in front and three behind, (B) Miohippus, an Oligocene three-toed horse, (C) Merychippus, a late Miocene form with reduced lateral toes, and (D) Equus. (From Vertebrate Paleontology by Alfred Sherwood Romer published by The University of Chicago Press, copyright © 1945, 1966 by The University of Chicago. All rights reserved. This material may be used and shared with the fair-use provisions of US copyright law, and it may be archived and redistributed in electronic form, provided that this entire notice, including copyright information, is carried and provided that the University of Chicago Press is notified and no fee is charged for access. Archiving, redistribution, or republication of this text on other terms, in any medium, requires both the consent of the authors and the University of Chicago Press.)
The significance of the fossil record of horses becomes clearer when it is compared with that of the other members of the order Perissodactyla ("odd-toed ungulates"). The fossil record of the extinct titanotheres is quite good (Fig. 7), and the earliest representatives of this group are very similar to "Eohippus" (Stanley, 1974; Mader, 1989). Likewise, the earliest members of the tapirs and rhinos were very "Eohippus"-like. Thus, the different perissodactyl groups can be traced back to a group of very similar small generalized ungulates (Radinsky, 1979; Prothero, et al., 1989; Prothero & Schoch, 1989) (Fig. 8). But this is not all; the most primitive ungulates (hoofed mammals) are the condylarths, which are assemblages of forms transitional in character between the insectivores and true ungulates (Fig. 9). Some genera and families of the condylarths had been previously assigned to the Insectivora, Carnivora, and even Primates (Romer, 1966). Thus, the farther you go back in the fossil record, the more difficult it is to place species in their "correct" higher taxonomic group. The boundaries of taxa become blurred.
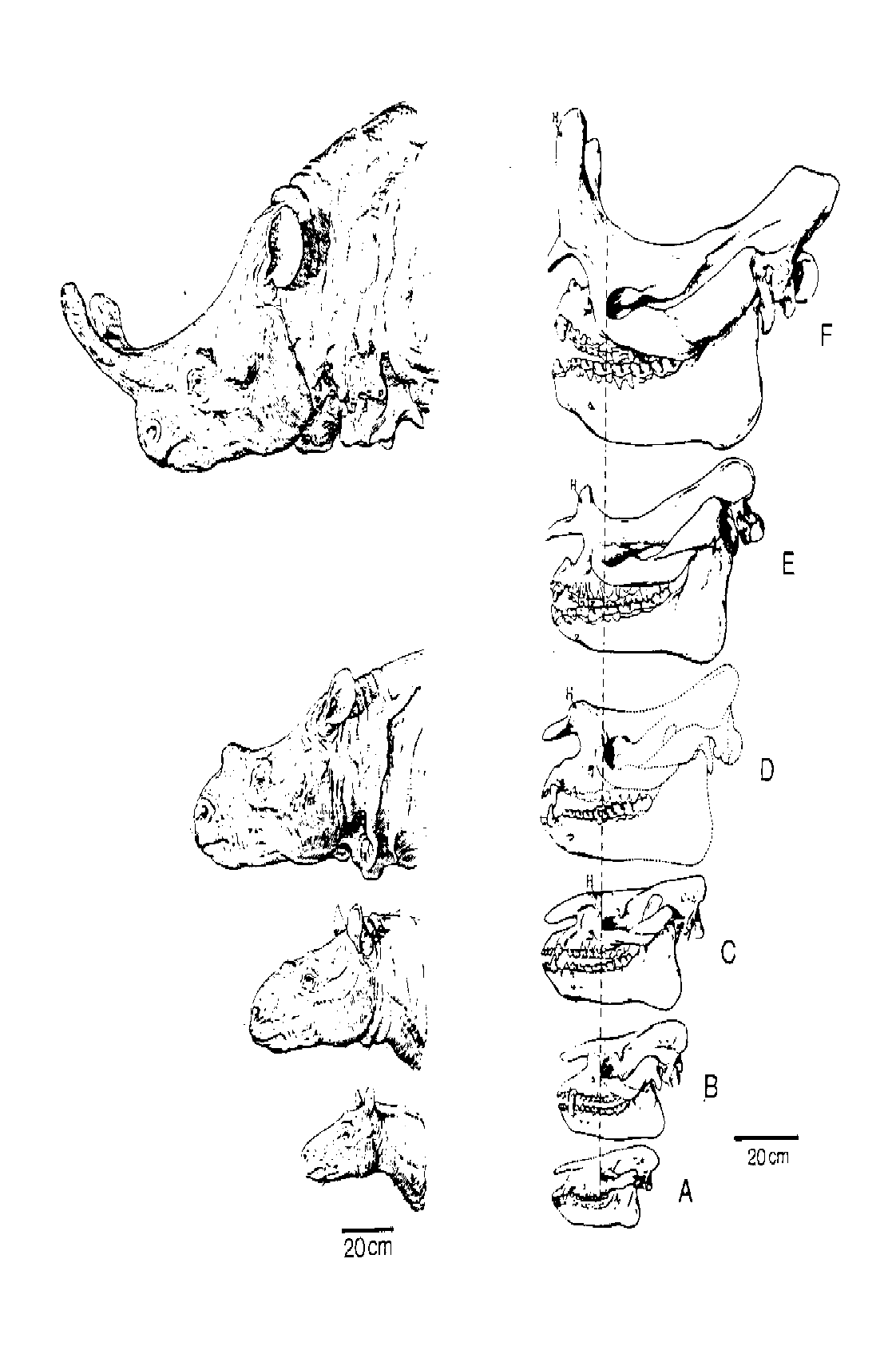
Figure 7. Stages in the evolution of the extinct perissodactyl family of the titanotheres. (A) Eotitanops (early Eocene), (B) Limnohyops (middle Eocene), (C) Manteoceras (middle Eocene), (D) Protitanotherium (late Eocene), (E) Brontotherium (early Oligocene), and (F) Brontotherium. (From Stanley [1974], reprinted with permission of the journal Evolution.)
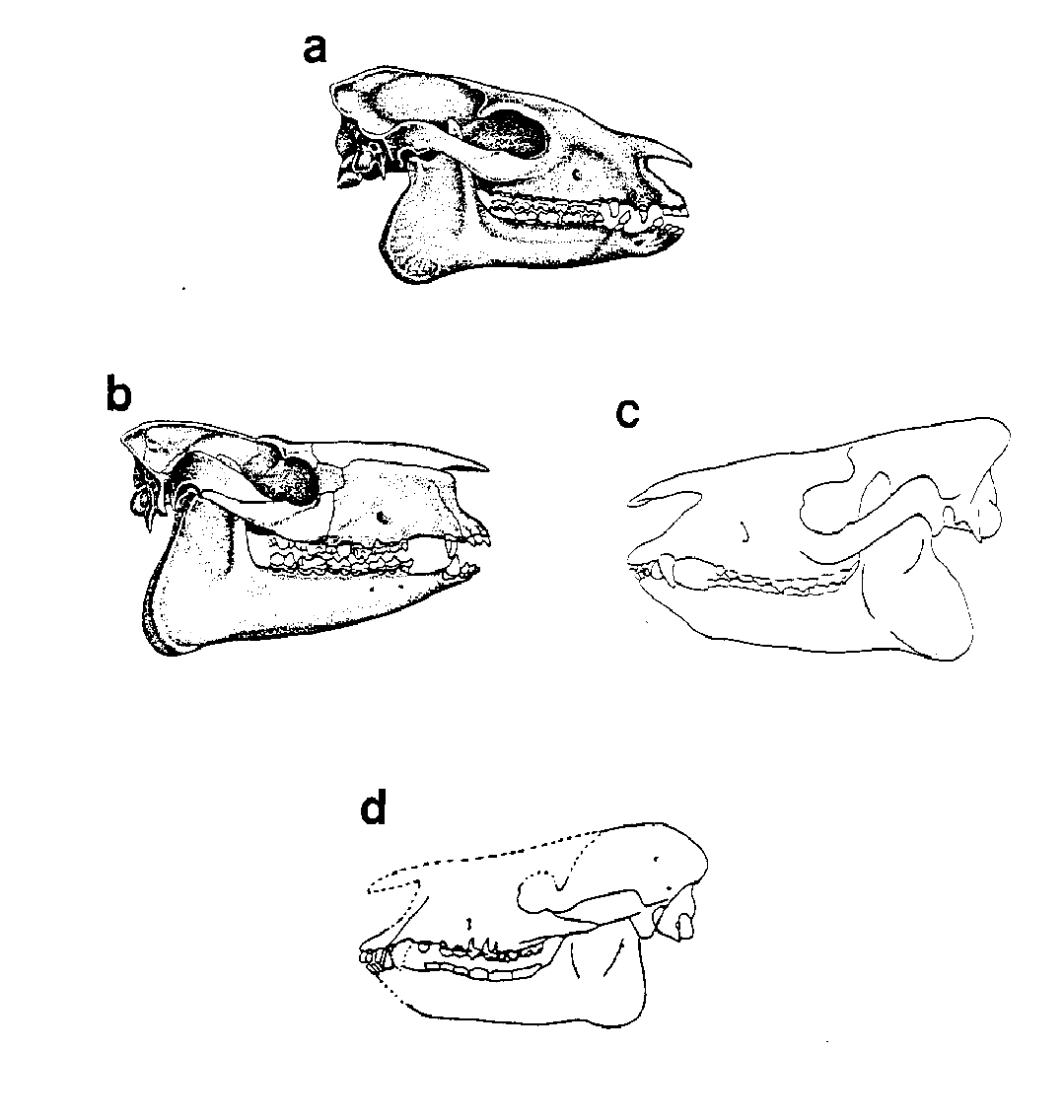
Figure 8. Comparison of the early members of four perissodactyl families. (A) Hyracotherium (Equoidea), (B) Hyrachyus (Rhinoceratoidea), (C) Heptodon ("Tapiroids"), (D) Eotitanops (Titanotheriomorpha). (A and B from Vertebrate Paleontology by Alfred Sherwood Romer published by The University of Chicago Press, copyright © 1945, 1966 by The University of Chicago. All rights reserved. This material may be used and shared with the fair-use provisions of US copyright law, and it may be archived and redistributed in electronic form, provided that this entire notice, including copyright information, is carried and provided that the University of Chicago Press is notified and no fee is charged for access. Archiving, redistribution, or republication of this text on other terms, in any medium, requires both the consent of the authors and the University of Chicago Press.) (C from Radinsky [1979], and D from Stanley [1974] both reprinted with permission of the journal Evolution.)

Figure 9. (A) The Eocene horse (Hyracotherium) and representatives of the condylarths, (B) Phenacodus (early Eocene) and (C) Mesonyx (middle Eocene). Note how very carnivore-like Mesonyx is although it possessed small hooves rather than claws and is classified with the ungulates. (From Vertebrate Paleontology by Alfred Sherwood Romer published by The University of Chicago Press, copyright © 1945, 1966 by The University of Chicago. All rights reserved. This material may be used and shared with the fair-use provisions of US copyright law, and it may be archived and redistributed in electronic form, provided that this entire notice, including copyright information, is carried and provided that the University of Chicago Press is notified and no fee is charged for access. Archiving, redistribution, or republication of this text on other terms, in any medium, requires both the consent of the authors and the University of Chicago Press.)
Moving further up the taxonomic hierarchy, the condylarths and primitive carnivores (creodonts, miacids) are very similar to each other in morphology (Fig. 9, 10), and some taxa have had their assignments to these orders changed. The Miacids in turn are very similar to the earliest representatives of the Families Canidae (dogs) and Mustelidae (weasels), both of Superfamily Arctoidea, and the Family Viverridae (civets) of the Superfamily Aeluroidea. As Romer (1966) states in Vertebrate Paleontology (p. 232), "Were we living at the beginning of the Oligocene, we should probably consider all these small carnivores as members of a single family." This statement also illustrates the point that the erection of a higher taxon is done in retrospect, after sufficient divergence has occurred to give particular traits significance.

Figure 10. Comparison of skulls of the early ungulates (condylarths) and carnivores. (A) The condylarth Phenacodus possessed large canines as well as cheek teeth partially adapted for herbivory. (B) The carnivore-like condylarth Mesonyx. The early Eocene creodonts (C) Oxyaena and (D) Sinopa were primitive carnivores apparently unrelated to any modern forms. (E) The Eocene Vulpavus is a representative of the miacids which probably was ancestral to all living carnivore groups. (From Vertebrate Paleontology by Alfred Sherwood Romer published by The University of Chicago Press, copyright © 1945, 1966 by The University of Chicago. All rights reserved. This material may be used and shared with the fair-use provisions of US copyright law, and it may be archived and redistributed in electronic form, provided that this entire notice, including copyright information, is carried and provided that the University of Chicago Press is notified and no fee is charged for access. Archiving, redistribution, or republication of this text on other terms, in any medium, requires both the consent of the authors and the University of Chicago Press.)
At the level of the class, the reptile/mammal transition is particularly well documented. Near the appearance of unquestioned mammals in the fossil record, a group of mammal-like reptiles called cynodonts included species that were exceptionally mammal-like in appearance (Hopson, 1994). In skeletal features the approach to the mammalian condition was almost complete (Fig. 11, 12). The following mammalian characteristics were possessed by advanced cynodonts: (1) enlarged temporal openings with the loss of the post-orbital bar, (2) absence of the pineal eye, (3) differentiation of teeth, with front nipping teeth, canines, and molar-like back teeth, (4) a secondary palate permitting respiration while chewing, (5) a double occipital condyle which enlarges the hole for the spinal cord, (6) absence of lumbar ribs (possibly related to the presence of a diaphragm), (7) a nearly erect stance, and (8) an enlarged dentary bone in the lower jaw with an extremely close approach to the mammalian jaw articulation. Furthermore, some workers argue persuasively that some mammal-like reptiles were endothermic (deRicqlés, 1974; Bakker, R.T., 1975; McNab, 1978). And a few exceptional fossils show evidence of glandular skin and horn (Hotton, 1991), features associated with the presence of hair.
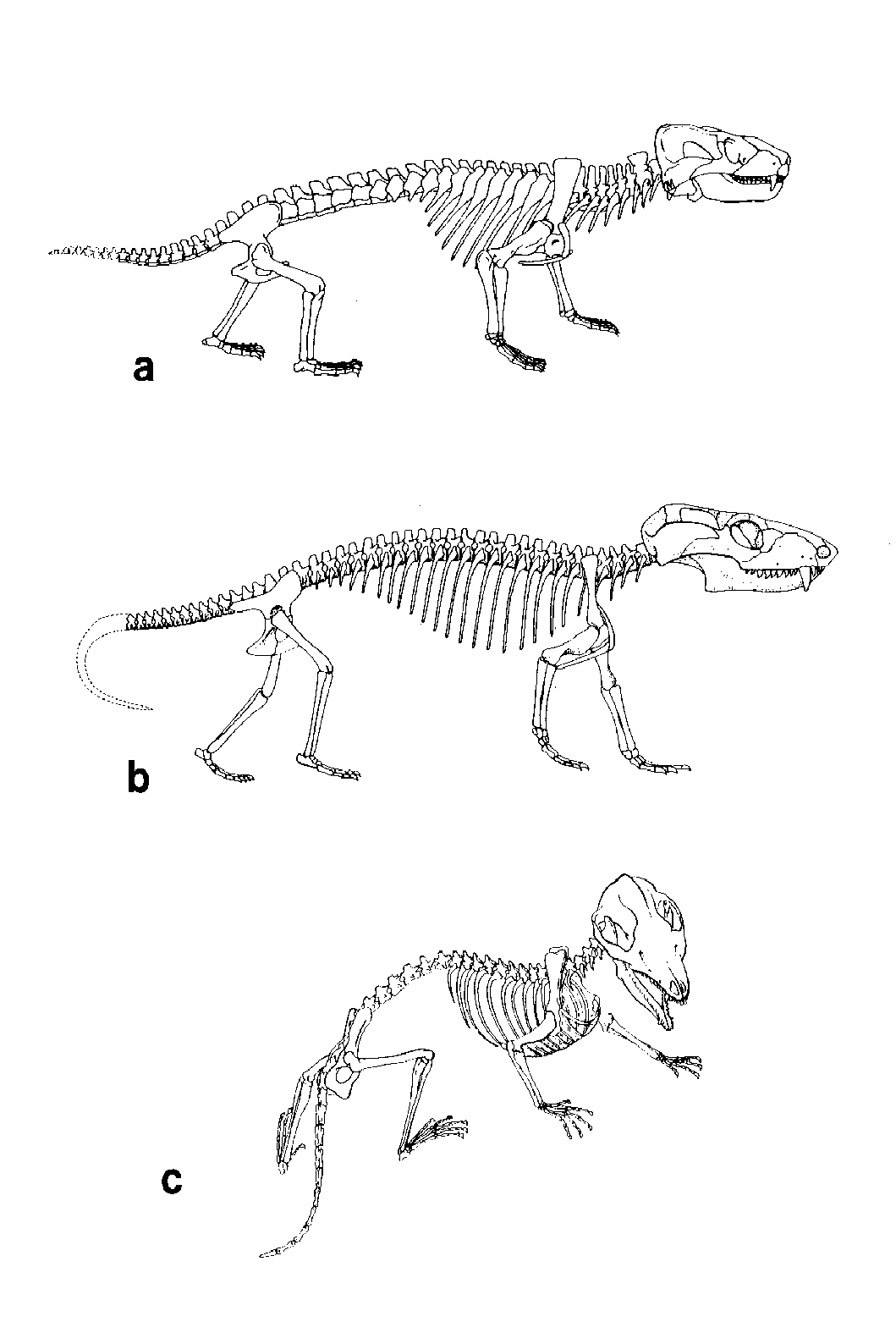
Figure 11. Reconstructed skeletons of cynodont (advanced mammal-like reptiles) and early mammals. (A) The early Triassic cynodont Thrinaxodon and (B) the advanced cynodont Probelesodon from the middle Triassic. Note the very mammal-like erect posture of these skeletons. (C) The early mammal Megazostrodon from the early Jurassic. (All reconstructions taken from Carroll [1988], A and C used by permission of Farish A. Jenkins, Jr., Museum of Comparative Zoology, Harvard University, and B used by permission of Arnold D. Lewis, Smithsonian Institution.)
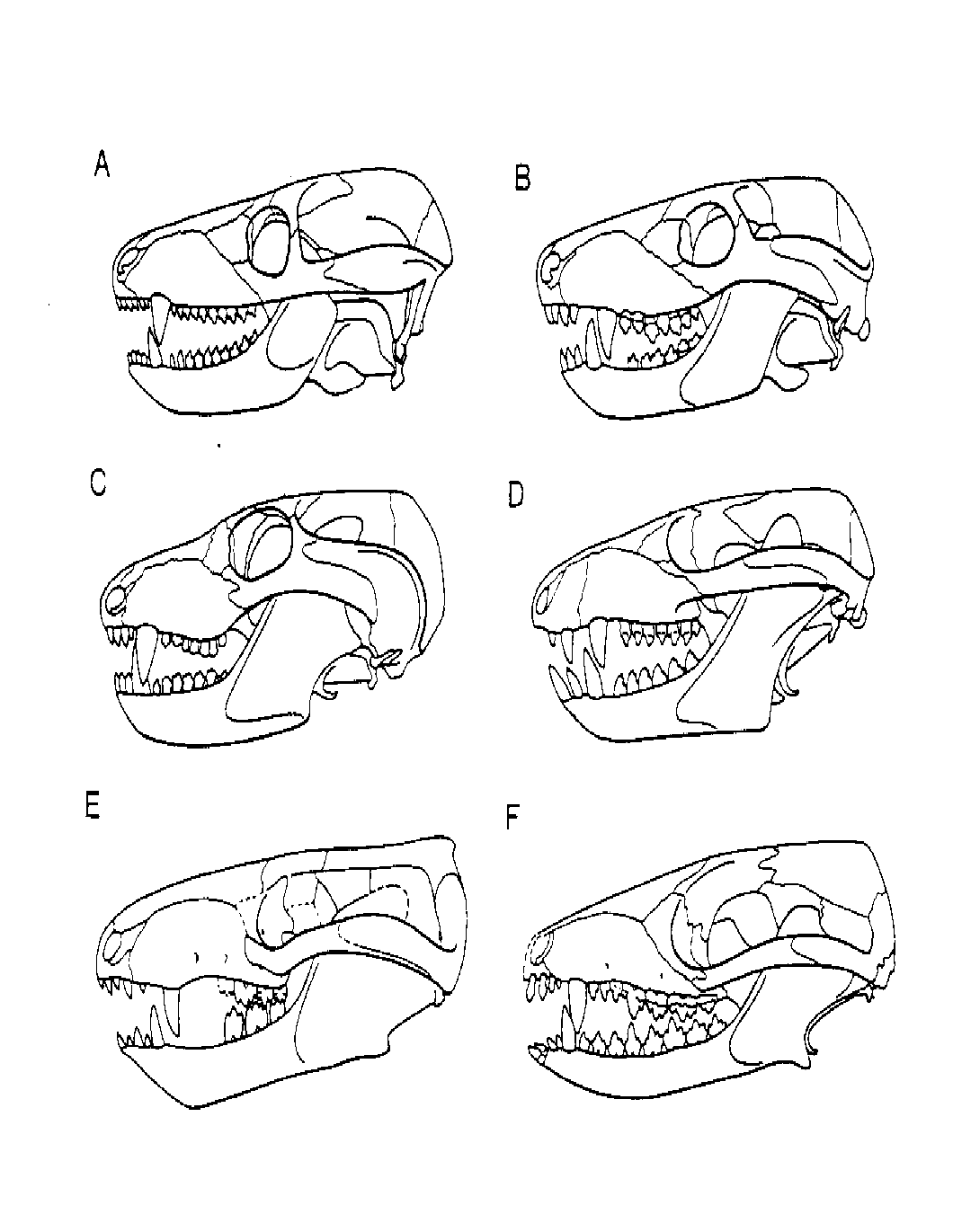
Figure 12. Comparison of the skulls of cynodonts and early mammals. The cynodont skulls are (A) the late Permian Procynosuchus; (B) the early Triassic Thrinaxodon; (C) the middle Triassic Probainognathus; and (D) the early Jurassic Pachygenelus. Note the differentiation of the teeth and the reduction in the bones at the back of the lower jaw. The early mammal skulls are (E) the early Jurassic Sinoconodon; and (F) the early Jurassic Morganucodon. (A through D from "Systematics of the nonmammalian Synapsida and implications for patterns of evolution in synapsids" by J.A. Hopson [1991], published in Origins of the Higher Groups of Tetrapods: Controversy and Consensus edited by H.-P. Schultze and L. Trueb. Used by permission of the publisher, Cornell University Press. This material is not to be printed or otherwise used without permission.) (E and F from Hopson [1994] and used by permission of James A. Hopson.)
The complex of transitional fossil forms has created significant problems for the definition of the class Mammalia (Desui, 1991). For most workers, the establishment of a squamosal-dentary jaw articulation is considered one of the primary defining characters. The transition in jaw articulation from reptiles to mammals is particularly illustrative of the appearance of a class level morphologic character (Fig. 12). In reptiles, the lower jaw contains several bones, and the articular bone at the back of the jaw articulates with the quadrate bone of the skull. In mammals, the lower jaw has only one bone, the dentary, and it articulates with the squamosal bone of the skull. Within the cynodont lineage, the dentary bone becomes progressively larger and the other bones are reduced to nubs at the back. In one group of advanced cynodonts, the dentary bone has been brought nearly into contact with the squamosal, and in another, a secondary articulation exists between the surangular (another small bone at the back of the jaw) and squamosal (Hopson, 1991). The earliest known mammals, the morganucodonts, retain the vestigial lower jaw bones of the reptiles. These small bones still form a reduced, but functional, reptilian jaw joint medial to the new dentary-squamosal mammalian articulation. These reptilian jaw elements were subsequently detached completely from the jaw to become the mammalian middle ear (Crompton & Parker, 1978). Better intermediate character states could hardly be imagined!
As with most transitions between higher taxonomic categories, there is more than one lineage that possesses intermediate morphologies. Again, this is consistent with both the expectations of evolutionary theory, and the nature of the fossil record. The prediction would be for a bush of many lineages, many of which would be dead ends. Because of their objective to erect only monophyletic taxa (an ancestor is grouped with all of its descendants), some paleontologists have advocated including mammals with the advanced cynodonts, or even with the whole group of mammal-like reptiles, in a single higher taxon (Desui, 1991).
As in the case of the reptile-mammal transition, the distinctiveness of the classes also becomes blurred during the amphibian-reptile transition. The oldest known reptiles (Fig. 13) have been collected within the fossilized stumps of lycopod trees from the late Pennsylvanian in Nova Scotia (Carroll, 1970, 1991). Several groups of reptiliomorph amphibians occur near the appearance of these unquestioned reptiles. Some of these (the seymouriamorphs and diadectomorphs) were in fact previously regarded as reptiles (Carroll, 1988; Benton, 1991).

Figure 13. Skeleton and skull of the earliest known reptile Hylonomus from the early Pennsylvanian. Reptiliomorph amphibians placed in a group called the anthracosaurs converge closely on the reptiles in skeletal morphology (see reconstructions of the anthracosaur amphibians Bruktererpeton and Proterogyrinus in Carroll [1991]). (From Vertebrate Paleontology by Alfred Sherwood Romer published by The University of Chicago Press, copyright © 1945, 1966 by The University of Chicago. All rights reserved. This material may be used and shared with the fair-use provisions of US copyright law, and it may be archived and redistributed in electronic form, provided that this entire notice, including copyright information, is carried and provided that the University of Chicago Press is notified and no fee is charged for access. Archiving, redistribution, or republication of this text on other terms, in any medium, requires both the consent of the authors and the University of Chicago Press.)
Of special interest in the history of life are the morphological transitions associated with the major adaptive shifts from water to land, land to water, and land to air. These major changes in mode of life opened up tremendous new adaptive opportunities for animal life. While the fossil evidence for some of these transitions is minimal, for others exciting parts of the puzzle have been uncovered.
The transition from water to land was one of the most significant events in animal evolution. Recent paleontological and systematic work has shed new light on this transition (Fig. 14). The most primitive amphibian yet known is the late Devonian Ichthyostega, a tetrapod with a flattened skull and bearing a tail fin. The limbs were until recently poorly known, but new fossil evidence has come to light. The hand, previously unknown, shows that these amphibians possessed seven to eight digits. The limbs also had a very limited range of movement and the animal was not as well adapted to terrestrial locomotion as previously thought (Ahlberg & Milner, 1994). The rhipidistian fishes are widely considered to have given rise to the amphibians. One small group of late Devonian rhipidistians, the panderichthyids, appears to be closely related to the ichthyostegids (Schultze, 1991). These fishes have flattened skulls very similar to that of the early amphibians. In addition, the anal and dorsal fins are absent, and the tail is very similar to that of Ichthyostega (Vorobyeva & Schultze, 1991). The lobed pectoral and pelvic fins have bones that homologize with the limb bones of the tetrapods. Whether part of a single direct lineage or not, ichthyostegid amphibians and panderichthyid fishes are clearly transitional forms between class level taxa. The first known skull of a panderichthyid was in fact initially considered to be an amphibian (Vorobyeva & Schultze, 1991), again illustrating the taxonomic problems encountered during the appearance and early radiation of a new taxon.
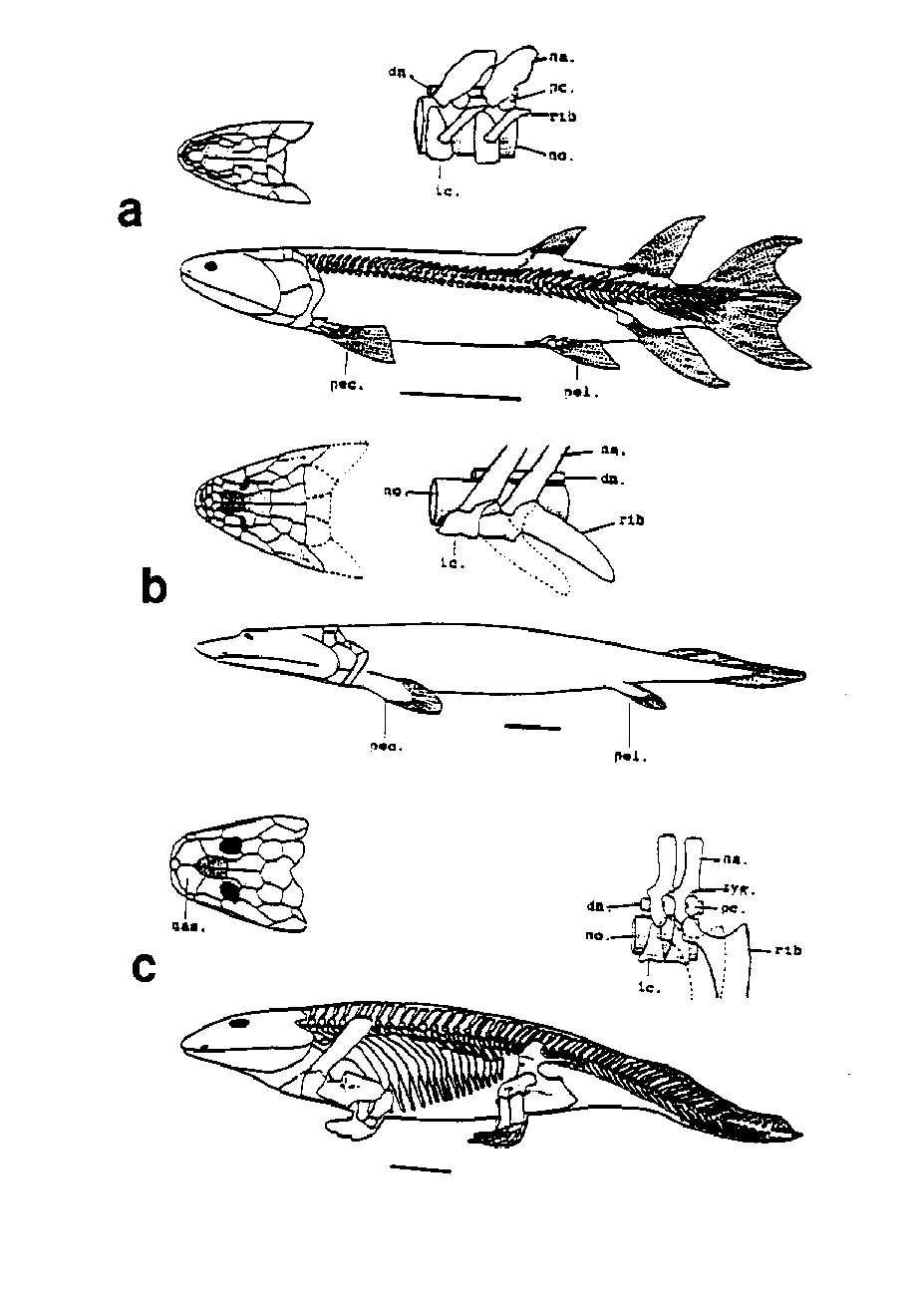
Figure 14. The transition from fish to amphibian illustrated by body form and skeletons, with details of skulls and vertebrae. (A) Osteolepiform fish Eusthenopteron; (B) panderichthyid fish Panderichthys; and (C) labyrinthodont amphibian Ichthyostega. (From Ahlberg & Milner [1994], reprinted with permission from Nature, copyright © 1994 Macmillan Magazines Limited, and from Per Ahlberg.)
Probably one of the most celebrated and mysterious transitions has been that of the origin of whales from a primitive condylarth (ungulate) ancestor. The earliest whales possessed skulls similar in many ways to those of a group of Eocene carnivorous condylarths called mesonycids. Until 1993 the earliest fossil whales were only known from partial skulls with no postcranial material. However, several very important transitional fossils from Pakistan have been described over the last several years (Gingerich, et al., 1993) and more discoveries are certain to follow. The geologically oldest included enough of the skeleton to reveal that this otter-sized whale had short front limbs and longer hind legs with large feet apparently used in swimming (Berta, 1994; Thewissen, et al., 1994). The second, somewhat younger species had shorter hind limbs indicating a trend toward reduction in limb size (Gingerich, et al., 1994). Whales apparently evolved in what is now Pakistan since all the known fossil material for earliest whales has been found in that geographic area. Because the evolution of new body plans is likely to occur in an isolated geographic area, the discovery of the fossil record of such transitions is dependent on the serendipitous sampling of the right locality.
The most famous of transitional fossils is the earliest known bird, Archeopteryx. Ostrum has described over 20 shared characteristics between Archeopteryx and coelurosaur theropods. Among these are: a theropod-like pelvis, the close similarities of the bones of the forelimbs including a swivel wrist joint, and the similarity of the hind limbs and feet with the presence of a reversed first toe (Hecht, et al., 1985; Dodson, 1985; Ostrom, 1994). The similarities of Archeopteryx to theropod dinosaurs such as Velociraptor and Deinonychus are especially strong, and a newly discovered dinosaur called Unenlagia has features of the limbs and pelvis that are the most bird-like yet known (Novas & Puerta, 1997). As interesting as the similarities with the theropods are, the differences between Archeopteryx and modern birds are also significant: it has a long bony tail, a sternum is absent, its vertebrae are not fused together over the pelvis to form a synsacrum, and air ducts are absent in its long bones. In most respects, Archeopteryx is more of a flying feathered dinosaur than a bird. In the last several years the discovery of new fossil birds from the Cretaceous has led to the erection of a whole new subclass of primitive birds called the enantiornithes (Chiappe, 1995). This new group includes several fossil species previously identified as theropod dinosaurs (e.g., Ornithomimus)! There are also some newly discovered fossils whose classification as theropod or bird is in dispute (Chiappe, 1995). The recent discovery in China of a theropod dinosaur with the possible preservation of fine feathers, even suggests that feathers may not be exclusively characteristic of birds (Morell, 1997). This again illustrates the taxonomic uncertainties that surround transitional forms.
From this brief survey of fossil vertebrates, it is clear that transitional forms between higher taxa are common features of the fossil record. The morphology of species within a higher taxonomic group becomes less divergent toward the point of origin of that group. Morphological diversity and disparity increase with time. In addition, transitional species possess mixtures of morphologic characters from different higher taxa often to the extent that their taxonomic assignment is uncertain. This pattern is obscured by taxonomy which gives a false impression of discontinuity.
The fossil record thus provides good evidence for the large-scale patterns and trends in evolutionary history. Recognizing its limitations, the fossil record appears to be consistent with the wide range of evolutionary mechanisms already proposed. Any wholesale abandonment of present paradigms would be very premature. Many critical gaps in our knowledge remain, but as evident from this review important discoveries are continually being made that intrigue, surprise, and enrich our understanding of the evolutionary history of life.
Ahlberg, P.E. and Milner, A.R., 1994, The origin and early diversification of tetrapods: Nature, vol. 368, p. 507-14.
Aldridge, R.J. and Purnell, M.A., 1996, The conodont controversies: Trends in Ecology and Evolution, vol. 11, p. 463-8.
Bakker, R.T., 1975, Dinosaur renaissance: Scientific American, vol. 232, p. 5878.
Benton, M.J., 1991, Amniote phylogeny: IN, H.-P. Schultze and L. Trueb, (eds.), Origins of the Higher Groups of Tetrapods: Controversy and Consensus, Comstock Publishing Associates, Ithaca, p. 317-30.
Berta, A., 1994, What is a whale?: Science, vol. 263, p. 180-1.
Briggs, D.E.G., Clarkson, E.N.K., and Aldridge, R.J., 1983, The conodont animal: Lethaia, vol. 16, p. 1-14.
Carroll, R.L., 1970, The earliest known reptiles: Yale Scientific Magazine, October issue, p. 16-23.
Carroll, R.L., 1988, Vertebrate Paleontology and Evolution: W.H. Freeman & Co., New York, 698 p.
Carroll, R.L., 1991, The origin of reptiles. IN, H.-P. Schultze and L. Trueb (eds.), Origins of the Higher Groups of Tetrapods: Controversy and Consensus, Comstock Publishing Associates, Ithaca, p. 331-53.
Chiappe, L.M., 1995, The first 85 million years of avian evolution: Nature, vol. 378, p. 349-55.
Crompton, A.W. and Parker, P., 1978, Evolution of the mammalian masticatory apparatus: American Scientist, vol. 66, p. 192-201.
Denton, M, 1985, Evolution: A Theory in Crisis: Adler & Adler, Bethesda, MD, 368 p.
deRicqlés, A.J., 1974, Evolution of endothermy: histological evidence: Evolutionary Theory, vol. 1, p. 51-80.
Desui, M., 1991, On the origins of mammals. IN, H.-P. Schultze and L. Trueb (eds.), Origins of the Higher Groups of Tetrapods: Controversy and Consensus, Comstock Publishing Associates, Ithaca, p. 570-97.
Dodson, P., 1985, International Archaeopteryx Conference: Journal of Vertebrate Paleontology, vol. 5, p. 177-9.
Eldredge, N. and Cracraft, J., 1980, Phylogenetic Patterns and the Evolutionary Process: Columbia University Press, New York, 349 p.
Erwin, D.H., 1993, The Great Paleozoic Crisis: Life and Death in the Permian: Columbia University Press, New York, 327 p.
Evander, R.L., 1989, Phylogeny of the family Equidae. IN, D.R. Prothero and R.M. Schoch (eds.), The Evolution of the Perissodactyls: Oxford University Press, New York, p. 109-27.
Gingerich, P.D., 1976, Paleontology and phylogeny: Patterns of evolution at the species level in early Tertiary mammals: American Journal of Science, vol. 276, p. 1-28.
Gingerich, P.D., Raza, S.M., Arif, M., Anwar, M., and Zhou, X., 1993, Partial skeletons of Indocetus ramani (Mammalia, Cetacea) from the lower middle Eocene Domanda Shale in the Sulaiman Range of Punjab (Pakistan): Contributions from the Museum of Paleontology, University of Michigan, vol. 28, p. 393-416.
Gingerich, P.D., Raza, S.M., Arif, M., Anwar, M., and Zhou, X., 1994, New whale from the Eocene of Pakistan and the origin of cetacean swimming: Nature, vol. 368, p. 844-7.
Hecht, M.K., Ostrom, J.H., Viohl, G., and Wellnhofer, P. (eds.), 1985, The Beginnings of Birds: Proceeding of the International Archaeopteryx Conference, Eichstatt, 1984: Bronner & Daentler, Eichstatt.
Hopson, J.A., 1991, Systematics of the nonmammalian Synapsida and implications for patterns of evolution in synapsids. IN, H.-P. Schultze and L. Trueb (eds.), Origins of the Higher Groups of Tetrapods: Controversy and Consensus, Comstock Publishing Associates, Ithaca, p. 635-93.
Hopson, J.A., 1994, Synapsid evolution and the radiation of non-eutherian mammals. IN, D.R. Prothero and R.M. Schoch (eds.), Major Features of Vertebrate Evolution, Short Courses in Paleontology, No. 7: Paleontological Society, Knoxville, p.190-219.
Hotton, N, III, 1991, The nature and diversity of synapsids: Prologue to the origin of mammals. IN, H.-P. Schultze and L. Trueb (eds.), Origins of the Higher Groups of Tetrapods: Controversy and Consensus, Comstock Publishing Associates, Ithaca, p.598-634.
Mader, B.J., 1989, The Brontotheriidae: A systematic revision and preliminary phylogeny of North American genera. IN, D.R. Prothero and R.M. Schoch (eds.), The Evolution of the Perissodactyls: Oxford University Press, New York, p. 109-27.
McFadden, B.J., 1988, Horses, the fossil record, and evolution: Evolutionary Biology, vol. 22, p. 131-58.
McFadden, B.J., 1992, Fossil Horses: Systematics, Paleobiology, and Evolution of the Family Equidae: Cambridge University Press, Cambridge, 369 p.
McNab, B.K., 1978, The evolution of endothermy in the phylogeny of mammals: American Naturalist, vol. 112, p. 1-21.
Mikulic, D.G., Briggs, D.E.G., and Kluessendorf, J., 1985, A Silurian soft-bodied fauna: Science, vol. 228, p. 715-7.
Morell, V., 1997, The origin of birds: the dinosaur debate: Audubon, vol. 99, no. 2, p. 36-45.
Norman, D., 1985, The Illustrated Encyclopedia of Dinosaurs: Crescent Books, New York, 208 p.
Norman, D., 1994, Prehistoric Life: The Rise of the Vertebrates: Macmillan, New York, 246 p.
Novas, F.E. and Puerta, P.F., 1997, New evidence concerning avian origins from the Late Cretaceous of Patagonia: Nature, vol. 387, p. 390-2.
Ostrum, J.H., 1979, Bird flight: How did it begin?: American Scientist, vol. 67, p. 46-56.
Ostrum, J.H., 1994, On the origin of birds and of avian flight. IN, D.R. Prothero and R.M. Schoch (eds.), Major Features of Vertebrate Evolution, Short Courses in Paleontology, No. 7: Paleontological Society, Knoxville, p.160-77.
Prothero, D.R., Guerin, C., and Manning E., 1989, The history of the Rhinocerotoidea. IN, D.R. Prothero and R.M. Schoch (eds.), The Evolution of the Perissodactyls: Oxford University Press, New York, p.321-40.
Prothero, D.R. and Schoch, R.M., 1989, Origin and evolution of the Perissodactyla: Summary and synthesis: IN, D.R. Prothero and R.M. Schoch (eds.), The Evolution of the Perissodactyls: Oxford University Press, New York, p.504-29.
Radinsky, L.B., 1979, The early evolution of the Perissodactyla: Evolution, vol. 23, p. 308-28.
Romer, A.S., 1966, Vertebrate Paleontology: University of Chicago
Press, Chicago,
468 p.
Schoch, R.M., 1986, Phylogeny Reconstruction in Paleontology: Van Nostrand Reinhold Company, New York, 351p.
Schultze, H.-P., 1991, A comparison of controversial hypotheses on the origin of tetrapods. IN, H.-P. Schultze and L. Trueb (eds.), Origins of the Higher Groups of Tetrapods: Controversy and Consensus, Comstock Publishing Associates, Ithaca, p. 29-67.
Stanley, S.M., 1974, Relative growth of the titanothere horn: a new approach to an old problem: Evolution, vol. 28, p. 447-57.
Stanley, S.M., 1979, Macroevolution: Pattern and Process: WH. Freeman & Company, San Francisco, 332 p.
Thewissen, J.G.M., Hussain, S.T., and Arif, M., 1994, Fossil evidence for the origin of aquatic locomotion in archaeocete whales: Science, vol. 263, p. 210-2.
Valentine, J.W., 1992, The macroevolution of phyla. IN, J.H. Lipps and P.W. Signor (eds.), Origin and Early Evolution of the Metazoa: Plenum Press, New York, p. 525-53.
Vorobyeva, E. and Schultze, H.-P., 1991, Description and systematics of panderichthyid fishes with comments on their relationship to tetrapods. IN, H.-P. Schultze and L. Trueb (eds.), Origins of the Higher Groups of Tetrapods: Controversy and Consensus, Comstock Publishing Associates, Ithaca, p. 68-109.