On Human Origins: A Survey
John A. Bloom, Ph.D., Ph.D. Biola
University
It is the theory which decides what we can observe. -- Albert Einstein1
Until
about 150 years ago, the subject of human origins was almost exclusively the
domain of religion. Revelation
from the gods passed down through cultural tradition told man how he came to be
and about his place in the Universe.
Since the time of Darwin, however, material from the natural world
itself has been discovered that allows the sciences to address the topic as
well. Over the past thirty years,
intensive field work in Africa as well as striking advances in biochemistry
have fostered a virtual explosion of new data which now must be incorporated into
our models about origins.
Integrating
a multidisciplinary database into a coherent, unified model is a difficult
task. This becomes especially
challenging when the issue involves our own origin, since here we cannot be
neutral observers. The meaning and
value which we assign to life are linked to the origins model we espouse. Since our significance depends on our
pedigree, those who desire there to be no supreme authority over their lives
will favor a different origins model than those who seek to glorify their
divine Creator with their lives.
Unfortunately,
most of the contact which nonspecialists have with human origins comes through
the mass media, where a secular-humanistic bias seems to pervade the scientific
content.2 For the person who seeks to integrate
this data with conservative Christian theology, it is difficult to judge from
these sources which new discoveries are important and need to be incorporated
into the synthesis, and which are but the artifacts of an overly zealous grant
proposal or hidden personal agenda.
The
goal of this article is to provide a neutral overview of some of the current
data and issues in the human origins debate. Without getting buried by details, we will attempt to give
the nonspecialist some feeling for the background to key scientific findings
that bear on a conservative Christian reading of Genesis. After providing some historical
background and outlining the relevant fossil and artifact evidence, we will
consider some criticisms of this data.
We will then review recent biochemical research and its contributions to
this issue. Since space does not
permit us to construct a fully integrated biblical-scientific model from the
data, we will focus the discussion on only one apparent clash between the
biblical and scientific models, that regarding the special creation of the
first man. Our proposed resolution
will serve as an example of the integrative tension and challenges that are
present in this field today.
The
first major treatises to argue for human origins from a solely naturalistic
outlook were T. H. Huxley's Man's Place in Nature (1863) and Charles Darwin's The Descent of Man (1871).
Although Neandertal3 fossils had been recovered on the
Continent starting in 1856, they were considered to be irrelevant to the
discussion because at that time their unique features were viewed as racial
variation, not as non- or pre-human differences of evolutionary import4. Thus Huxley’s and Darwin's
arguments hinged on man's anatomical homology with modern apes and lower
primates, not on any fossil data.
The
quest for fossil evidence of a common ancestor for apes and man (popularly
known as the “missing link”) was taken up by Eugène Dubois,
a French anatomist who was inspired by the works of Ernst Haeckel. Haeckel, a leading German zoologist in
the 1880's and a strong proponent of macroevolution,5 proposed that the
remains of "Ape-like Men" (to which he assigned the name Pithecanthropus) might be found on the continent of Lemuria [an
ancient land mass proposed by nineteenth-century geologists that was believed
to have sunk beneath the Indian Ocean].
Dubois took a medical position with the Dutch East India Army stationed
in Java (then a colony of Holland) in order to be as close to the hypothetical
homeland of the missing link as possible and proceeded to excavate in the
fossil beds there.
After
several years of work, Dubois had collected a primitive skull cap, tooth, femur
(thigh bone), and several other bone fragments. The small brain size evidenced by the skull cap coupled with
the modern appearance of the femur led him to the conclusion that he had found
a transitional ape-like man that could walk upright; hence in 1893 he named his
find Pithecanthropus erectus.6 Because the evidence was fragmentary
and it was not certain that all of the bones came from a single individual,
Dubois' conclusion was not well received by most of the scientific community
for many years.7
Unfortunately,
Charles Dawson's discovery of the Piltdown Man in 1913 did not meet with the
same skepticism because the find's combination of large brain size and ape-like
jaw fit better with the preconceptions of the day.8 Meanwhile, interest in Asia picked up
as other researchers found hominid fossils and tools in Zhoukoudian, China, and
were proposing a missing link in the form of the "Peking Man."9
In
1925 the search for early man turned to Africa (the homeland posited by Darwin
because it was the habitat of the modern ape and chimpanzee) when Raymond Dart
from South Africa reported the discovery of a fossilized skull of a very
primitive young hominid, whom he named Australopithecus africanus ("southern ape from Africa").10 Informally called the "Taung
baby," its relation to mankind was rejected11 for over two
decades, until Robert Broom12 and others recovered many more
australopithecine fossils. Early
twentieth-century scientific models and preconceptions about human evolution
then shifted to accommodate this mounting African evidence.
Louis
Leakey faced the same pro-Asian bias when he began work in East Africa in
1931. Years of effort at the now
famous Olduvai Gorge in Tanzania as well as sites in Kenya yielded the Zinjanthropus
boisei or "Nutcracker Man"
in 195913 and the first Homo habilis ("Handy Man") fossil in 1961,14 thus confirming the African-homeland
hypothesis in many people's minds.
Over
the past thirty years, the research efforts of paleoanthropologists have
increased in both quantity and quality.
It was estimated recently that we now have recovered the remains of at
least 275 Neandertal individuals from more than 70 sites,15
and the total number of known hominid fossil individuals exceeds six thousand.16 Consequently, rather than continuing to
present these results in the order of their discovery, it is much more logical
to summarize the material by the age of the fossil and its features.
II.
Generally Accepted Hominid Categories
Australopithecus
afarensis
The
most famous example of the species Australopithecus afarensis (“southern ape from Afar”) is called
"Lucy," discovered in 1974 by Donald Johanson at the Hadar Formation
in Ethiopia.17
Although similar specimens were found later at other east African sites,18
Lucy is unique because about 40% of her skeleton was recovered. Finding such a large fraction of bones
from a single individual is striking, given the great age of the fossils (dated
to about 3.2 million years ago [mya]).19 20 Standing between 3.5 to 5 feet tall, A.
afarensis was thick-boned with
proportionally shorter legs and longer arms than modern humans. From skeletal remains it was postulated
that this species was fully bipedal, and the later discovery at Laetoli of a
3.7 million-year-old extensive track of hominid footprints made by three
bipedal individuals21 appears to bear out this hypothesis
dramatically. Despite her advanced
pelvis and mode of walking, Lucy's skull is very similar to a chimp's, with a
small brain (400 cc average), and large teeth.22
Other
Australopithecine species.
In
the fall of 1994, the discovery of bone fragments from a more primitive
australopithecine than A. afarensis
was announced.23
Called A. ramidus, these
fossils are dated at 4.4 mya (over a million years older than Lucy), and should
help show how the transition was made from early arboreal monkey-like primates
to walking hominids.
Australopithecine
hominids that appear in the record after Lucy are generally classed into at
least three groups, A. africanus, A. boisei and A. robustus, based on their body size and cranial features. They have larger stature than afarensis, are also bipedal, and differ markedly from each
other in their skull structure, probably due to the influence of diet.24 Since only africanus has dentition that is comparable with that found the
later homo series, it is the only
late australopithecine that some consider to be a direct ancestor to modern
man.
Homo
habilis
Homo
habilis is the name given to a group
of African fossils that are believed to have used the first primitive stone
tools, called the Oldowan tradition.25 Spanning a time range from 2.3 to 1.5 mya, H. habilis was clearly bipedal, thin boned, and had a larger
brain size (670 cc) than A. afarensis. One of the best H. habilis specimens is the nearly complete skull (KNM-ER-1470)
found by Richard Leakey.26
However, recent new finds by Johanson (OH 62) suggest that H. habilis was not significantly taller than A. afarensis and had a similar short leg / long arm structure.27 This apparent mix of change and
stability in H. habilis (a more
human-like skull with a larger brain and smaller teeth than afarensis is still combined with an ape-like limb structure)
makes it difficult to use in gradualist evolutionary schemes as a transitional
species between A. afarensis and H.
erectus. To remedy this problem, some suggest that this species
should not be regarded as a human ancestor (i.e., a member of the genus Homo) but rather as a distinct evolutionary branch.28 Because
there is considerable diversity in features among the specimens initially
included in the H. habilis taxon,
the current trend is to reassign the specimens into other better-known groups
or divide them more finely-specified taxa.29 Thus Leakey’s KNM-ER-1470 specimen is reclassified as Homo
rudolfensis.
H.
habilis is the presumed occupant of
the earliest known (dated to 1.8 mya) living area: This 2,400-square-foot site discovered by Mary Leakey at
Olduvai Bed I contained over 4,000 artifacts and fossils. It provides direct
evidence of simple windbreaks or shelters and suggests that early hominids had
a nomadic hunter-gatherer lifestyle.30
Homo
erectus
Homo
erectus remains cover a period from
1.8 million to 300,000 years ago and are the first hominids to have a global
(Old World) distribution. The most
striking H. erectus find so far is
undoubtedly Richard Leakey’s “Turkana Boy,” the nickname
given to most of the skeleton of a large adolescent that dates to 1.6 mya
(KNM-WT-15000).31
As with habilus, the
current trend with erectus
specimens is to split them up rather than lump them into one taxon, so the
Turkana Boy now is listed as an “early African h. erectus” or h. ergaster. H. erectus manufactured the more advanced flaked stone tools of
the Acheulian tradition, were involved in big-game hunting, and had the
controlled use of fire. In many
skeletal features they appear essentially modern.32 The largest differences occur in the
skull, which is markedly flatter and thicker boned than modern humans. Moreover, the brow ridge above the eyes
is very pronounced, the jawbone lacks a protruding chin, and the teeth are
larger than moderns. The brain
size of H. erectus appears to
increase over time from 750 to 1,200 cc.
Since a brain size of greater than about 800cc in a light-boned hominid
places this species within the brain size range of modern man, some argue that H.
erectus is the first true modern man.33 Others are hesitant to lump H.
erectus with modern man solely on
this criteria, noting that the Acheulian toolkit is comparatively simple and
remained stable for over a million years, and thus does not reflect the
innovation and creativity that characterize modern human artifacts.34
The
global (old World) variation and migration patterns of the h erectus family have been the focus of study in recent
years. The distinctions between
Asian erectus (old popular names
being the “Peking” and “Java” man) and African erectus, combined with the very early date of some of the
Asian fossils,35 gave raise to speculations that these
regional groups may have arisen from a much earlier migration than previously
considered (i.e., a h habilus migration
instead of a h erectus one), or
that h erectus actually evolved in
Asia and migrated into Africa, a reversal of the dominant
“out-of-Africa” theory. However, a new erectus find in Africa is of comparable date to the Asian fossils and shows a
marked similarity to them, strengthening the single-species theory.36
Archaic
Homo sapiens
Early (or archaic)
Homo sapiens applies to a group of
fossils that share a mixture of H. erectus and modern features [A recent trend in naming this group is to avoid
the term “archaic” and to list them as Homo heidelbergensis (with variation)]. Most of these date from 300,000 to 100,000 years ago. In general the skeletal bones are less
robust than H. erectus while the
cranial capacity is larger, but the features are blurred enough that
considerable debate surrounds their classification. Many scholars feel that there is essentially a continuum
between late H. erectus and early
Homo sapiens both in features and
culture. For example, early
Homo sapiens tools are basically of
the Acheulian tradition, although some items (such as handaxes) apparently
become less popular and we begin to see some refinements in toolmaking
technique.37
Archaic
Homo sapiens sites attest to a
lifestyle of seasonal migration. A
shore site at Terra Amata in France dated to about 300,000 years ago yielded
large oval huts with a hearth at the center, large numbers of stone tools, and
possible evidence for body decoration.38
Neandertals
Neandertals are noted separately from archaic Homo sapiens because of their historical fame and the ongoing
debate regarding their classification.
Their bodies, teeth and jaws are very robust by modern standards, and
they share many of the cranial features found in H. erectus (above).
All
of the Neandertal remains that have been recovered to date are restricted
geographically to Western Europe and the Middle East. Their tools (called the Mousterian culture) are more
sophisticated than H. erectus and
the evidence that they buried their dead is now generally accepted. Evidence of animal trapping as an
alternative to hunting or scavenging is also found. Their first remains date back 80,000 years ago, and it
appears that they were rapidly replaced by modern humans in Europe around
30,000 years ago.
The
debate surrounding Neandertals involves their relationship to modern man,
because the data are ambiguous even for secular minds: Is their culture sophisticated enough
and anatomy compatible enough that they should be considered an intermediate
step between H. erectus and modern
man, or do they form a separate branch that has no recent ancestral
relationship? Did Neandertals and
modern humans occupy the same geographic areas at the same time or
interbreed? The answer which
secular scholars give to these questions depends heavily on the model for
recent human prehistory which they assume (see below).
Anatomically
modern humans
Anatomically
modern humans (AMH), as the name
implies, refers to hominid remains that are indistinguishable from modern man
(allowing for some racial variation).
They first appear in the fossil record at several sites in Africa that
are dated between 125,000 to 100,000 years ago. The earliest remains have modern cranial features (high
forehead, smaller teeth) but use the same tools as their archaic H. sapiens contemporaries.39 Although there are some new tantalizing clues from Blombos
cave in South Africa that AMH culture had elements of art and advanced
tool-making skills over 70,000 years ago,40 a distinct change occurs
globally in the AMH record about 40,000 years ago, when culture that all
researchers are comfortable in assigning to modern man suddenly appears around
the world.41 We find
ingenious weapons like boomerangs, an increased use of bone for tools, evidence
of higher population densities than at previous times, of long distance
migration to Australia and the New World, of trade or the transport of exotic
materials over considerable distances, and -- most strikingly -- of artistic
expression (sculpture, jewelry, painting, and music). In artistic expression, some see the first preserved clues
to the religious interests of early man.
David Lewis-Williams and Thomas Dowson propose that shamanistic
experiences explain the unusual geometric shapes seen in wall paintings that
date back as far as 27,000 BC.42
The
origin of language, certainly an essential hallmark of modern man, is
frustrating to evaluate since language leaves no clear trace in the
archaeological record until the advent of writing (around 3,000 BC). Seeking secondary clues, researchers
have investigated the physiology of fossil skulls for evidence of Broca's and
Wernicke's areas (the two principal speech centers in the brain) and for
evidence of the modern human larynx structure that allows a full variety of
speech sounds to be produced. No
consensus has been reached in these studies; some argue that H. habilis had Broca's area43 and thus could speak, and
others argue that even the late Neandertal did not have our modern larynx and
thus had a less efficient vocal repertoire.44 Still other scholars note that just because an ancient
hominid had the physical capacity for speech does not prove that any
sophisticated language was in fact used.
Working from the other direction, linguists have attempted to evaluate
the rate at which modern languages develop and diverge over time so that some
estimate back to a single original tongue can be made. Depending on the assumptions made, the
evolution time for our present language diversity appears to be between 50,000
to 100,000 years, an age that seems linked to anatomically modern humans.45
III.
Fossils: Summary and Appraisal
If
one imagines the above fossil and artifact data (cranial capacity, complexity
of tools, date found in the fossil record, etc.) to form a series of points on
a graph, the model of human origins which one espouses will affect how one
draws the line connecting the points.
Someone who believes that God intervened in the special creation of man
will place at least one discontinuity somewhere on the graph, while someone who assumes that man
shares a common ancestry with apes will arrange the data and connect the points
so as to show a smooth transition from early primitive forms to late modern
ones. The later scheme is the
approach of secular naturalists and is shown in the figure below.
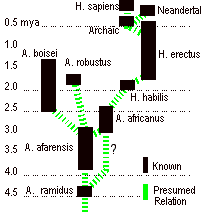 Prior
to 1979, it was generally assumed that competition for limited resources
permitted only one species to be the dominant hominid population in a region at
a given time. However, fossil
discoveries46 since then showed that A. boisei existed in Africa as a contemporary with A.
africanus, greatly weakening the
argument that africanus was
ancestral to robustus. In fact it appears that four or five
distinct hominid species coexisted in Africa around two mya, making the
evolutionary “tree” appear more like a bush during this
period. The new discovery of ramidus as the earliest known hominid replaces afarensis at the trunk of the tree, and reopens the debate
whether afarensis is really the
last common ancestor to the australopithecine and homo lines, as Johanson
proposed but Richard Leakey doubts.47 The recent discovery of K platyops, a contemporary hominid with afarensis, raises the possibility that the
“bushiness” seen at two mya extends all the way down, with the
result that we cannot discern a single, unambiguous ancestral lineage nor an
obvious “last common ancestor” with the primates.
Prior
to 1979, it was generally assumed that competition for limited resources
permitted only one species to be the dominant hominid population in a region at
a given time. However, fossil
discoveries46 since then showed that A. boisei existed in Africa as a contemporary with A.
africanus, greatly weakening the
argument that africanus was
ancestral to robustus. In fact it appears that four or five
distinct hominid species coexisted in Africa around two mya, making the
evolutionary “tree” appear more like a bush during this
period. The new discovery of ramidus as the earliest known hominid replaces afarensis at the trunk of the tree, and reopens the debate
whether afarensis is really the
last common ancestor to the australopithecine and homo lines, as Johanson
proposed but Richard Leakey doubts.47 The recent discovery of K platyops, a contemporary hominid with afarensis, raises the possibility that the
“bushiness” seen at two mya extends all the way down, with the
result that we cannot discern a single, unambiguous ancestral lineage nor an
obvious “last common ancestor” with the primates.
A
significant difficulty in evaluating the recent fossil data is that we are too
close to the personalities and biases of the discoverers. Paleoanthropologists, being human, can
be myopic about the overall significance of their own work: After all, everyone prefers to discover
something new rather than find the seventeenth specimen of an already
well-known species. Consequently
it is often the next generation of scholars who are more objective at
integrating data into a coherent scheme than the pioneers who made the original
discoveries. Bearing this in mind,
it is not unreasonable to find that scholars questioned the validity of a given
species, such as H. habilis, and
asked for a review of the specimens assigned to this taxon.48 It is even possible this taxon may
disappear because its fossils can be better assigned (on the basis of brain
size, limb-length ratios, and possible tool usage) to the better-characterized A.
africanus and H. erectus classes.
However, such a careful assessment is best done by those who have direct
access to the fossils.
As
noted above, once H. erectus
appears, the rest of human prehistory appears to be a continuum (if one accepts
evolutionary presuppositions).
There is some gradual change in erectus cranial features from early to late periods, but then
a blend of modern and erectus
features appear in early H. sapiens
and Neandertals just prior to and contemporaneous with fully modern forms.49 Some specimens exhibit such a mosaic of
features that a precise label assignment cannot be made. This makes it difficult to deduce when,
where, and how modern man appeared.
Lubenow,
a young-earth creationist who feels the earth is only a few thousand years old,
reviews the above fossil data and concludes that the diversity in the entire Homo taxon reflects only racial differences, not an
evolutionary or species development.50 Other researchers (both secular and Christian) consider the
dating information to be generally reliable, and have proposed other models
that take into account the time intervals and diversity observed. The key differences between these models
depend on their assumptions about the level and type of interaction among the
global H. erectus population.
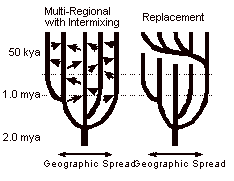 Following
the early intercontinental distribution of H. erectus around 1.5 mya, the multi-regional model proposes that this species globally evolved into
modern H. sapiens because a
sufficient level of interbreeding throughout the dispersed population was
maintained over this long time period.
Proponents for this view argue that the similarity of ancient and modern
skulls found in a given location (particularly China, Australia and the Far
East) implies that the hominid population in that geographic area has been
stable for hundreds of thousands of years.51 However, many doubt that H. erectus populations intermingled over thousands of miles at a
sufficient level to keep the advance to H. sapiens globally synchronized. On the other hand, the replacement model proposes that all modern populations result from a
recent global migration of mankind from a single locale that replaced any
earlier hominids (such as Neandertals) whenever they came in contact.52 Total replacement proponents are quick
to point out that "Rambo-like invasion" characterizations of their
model are inaccurate; a simple subsistence advantage that reduces mortality by
two percent in the new population will enable it to replace the older group
without violence in less than one thousand years.53 Moreover, in some geographical areas
there may have been nothing to replace: Earlier hominid populations may have
died out or been reduced to minimal levels by climate change or disease before
the immigrants arrived. Of course,
some hybrid of these two models is possible, that combines one or more late,
major waves of migration with some intermixing with older native populations.
Following
the early intercontinental distribution of H. erectus around 1.5 mya, the multi-regional model proposes that this species globally evolved into
modern H. sapiens because a
sufficient level of interbreeding throughout the dispersed population was
maintained over this long time period.
Proponents for this view argue that the similarity of ancient and modern
skulls found in a given location (particularly China, Australia and the Far
East) implies that the hominid population in that geographic area has been
stable for hundreds of thousands of years.51 However, many doubt that H. erectus populations intermingled over thousands of miles at a
sufficient level to keep the advance to H. sapiens globally synchronized. On the other hand, the replacement model proposes that all modern populations result from a
recent global migration of mankind from a single locale that replaced any
earlier hominids (such as Neandertals) whenever they came in contact.52 Total replacement proponents are quick
to point out that "Rambo-like invasion" characterizations of their
model are inaccurate; a simple subsistence advantage that reduces mortality by
two percent in the new population will enable it to replace the older group
without violence in less than one thousand years.53 Moreover, in some geographical areas
there may have been nothing to replace: Earlier hominid populations may have
died out or been reduced to minimal levels by climate change or disease before
the immigrants arrived. Of course,
some hybrid of these two models is possible, that combines one or more late,
major waves of migration with some intermixing with older native populations.
These
two models view the origin of advanced culture and language quite
differently: In the multi-regional
model, culture and language foster isolated pockets of hominids that intermix
gradually over time. The
replacement model typically sees culture and language as the competitive edges
aiding the global expansion of one advanced hominid from out of Africa (at
least this is where the oldest known fossils are found) sometime within the
past 100,000 years.
We
should note in passing that either of these secular models is compatible with a
special creation of mankind, either by placing a Homo erectus Adam at the root of the multi-regional model, or by
placing an anatomically modern human
Adam as the source of the replacing population. There are advantages and drawbacks to either approach, but
space does not permit a full analysis of both models. Consequently, we will limit our discussion to the hypothesis
the author favors, that of a recent modern Adam.
IV.
Biochemistry: The New Database
Besides
offering new tools for fossil analysis, other scientific disciplines are
finding new data sources that contribute to the origins question. The most significant of these involves
the realization that DNA, the chemical in the nucleus of every cell which
stores the information necessary to manufacture the proteins, cells, organs,
and structure of the entire human body, also contains clues about the
biological history of man.
Biochemists are now able to compare the DNA and protein sequences of
modern animals, and on the basis of the small differences found, they can
estimate the chemical similarity of man and the lower primates. The similarity appears to be very
high: DNA differs by about 2%, and
proteins by about 0.3% between human and chimpanzee, the closest lower
primate. Based on this small
difference and on estimated rates of sequence variation over time, it has been
deduced that a hypothetical last common ancestor to man and African apes
existed as recently as five to seven million years ago.54
Another
valuable biochemical insight involves the analysis of mitochondrial DNA from a
world-wide sample of people. This
form of non-nuclear DNA, which is only inherited from the mother, appears to
show that all living humans are more closely related to one another than the
multi-regional model of human evolution suggested. In fact, the mitochondrial DNA similarity is so high that
the initial researchers inferred that the female ancestor common to all modern
humans lived as recently as 90,000 to 180,000 years ago.55 However, this recent-date assignment
and its interpretation as a single individual has come under strong attack and
may need revision in light of statistical limitations inherent in the analysis.56 Nevertheless the data clearly show that
all humans are very closely related to each other. Studies of chimpanzees and apes show that much more genetic
diversity exists within each of
these primate species than exists within the human family. To many
this implies that a “pinch” or “bottleneck” occurred in
the human population in the recent past.57
Technological
improvements in the study of nuclear DNA now permit researchers to progress
beyond bulk comparisons of similarity (such as the 2% figure noted above) and
undertake a detailed comparative study of specific sequences. In some DNA regions, especially those
that carry the information for allowing the human immune system to distinguish
friend from foe (called the major histocompatability [MHC] genes), considerable
patterns of variation are observed between individuals. These patterns are used to form the
equivalent of molecular fingerprints on the cell surface, and their variability
is so high that it is very unlikely that any two people will have identical
cellular fingerprints. While this
variability made organ transplants very difficult before the advent of
immunosuppressant drugs, the benefit of this personalized cellular ID system is
that it is extremely difficult for bacteria and parasites to invade the body.
Research
has been undertaken to compare the MHC-gene patterns in humans with those of
other primates. These transspecies
polymorphism studies found that some patterns are more similar between species than within species.
The researchers deduce from this that the number of similar patterns
between species must be proportional to the number of common ancestors that
both species shared. For the sake
of illustration, let us say that there are ten patterns whose sequence is much
more similar between humans and chimpanzees than it is between any other
human-human or chimp-chimp combination.
We also know from our genetic studies that each individual can carry
only one of these patterns. The
fact that these ten human-chimp patterns exist in common today implies that at
least ten different individuals, each carrying one of the unique patterns,
comprised the group that formed the last common ancestors to both the human and
chimpanzee. In other words, the
number of sequence patterns which two species have in common is proportional to
the genetic diversity (i.e., group size) of the last common ancestors to both
species.58 Using this
reasoning with the currently known number of similar patterns between humans
and chimps allows the secular researchers to assert boldly that "MHC
polymorphism categorically rules out the possibility that modern human
populations are derived from a single individual -- an Eve."59
As
with any conclusion, the assumptions and data which lead to it need to be
carefully examined. For example,
the forces that give rise to the diversity seen in the MHC genes are poorly
understood. Similar patterns
between humans and chimpanzees may result because both need this specific
pattern to ward off a common parasite.
Consequently the MHC genes show a common “history” because
both populations (or population subgroups) have successfully -- but independently
-- fought off the same enemy. A
common history of exposure to pathogens is a far cry from common ancestry. It remains to be shown that the common
chimpanzee-human patterns arose without any external selection pressure. Otherwise, the data do not distinguish
a common pathogen or other convergent pressure from a common ancestor.
Furthermore,
continued transspecies polymorphism studies using larger population samples may
find too many similar human-chimp
patterns. Using the above logic,
this would make the required number of founding individuals impossibly large
and create a conflict with a fundamental assumption in evolutionary population
genetics: evolutionary change requires small populations. If the group is too large, genetic
novelties will not statistically dominate. As the human genome is better understood, the total number
of common patterns found between species may require an unreasonably large
founding population, implying that this transspecies similarity has other than
ancestral origin.
Lastly,
as the attacks on early mitochondrial-DNA conclusions have shown, the computer
programs used to deduce genetic similarity between variant sequences are not as
unbiased as previously thought.
They are affected by the order in which the data are entered and often
do not converge to a single obvious solution.60 Given the infancy of genetic research
and analysis tools, categorical conclusions are best avoided.
Just
as the mitochondrial DNA in a woman’s egg is passed on to all of her
descendants and does not mix with any genetic material from the man, the male Y
chromosome is passed on to all male descendants and does not mix with any
genetic information from the woman.
Consequently there is intense interest in studying the Y chromosome to
see if it shows the same very low variation that the mitochondrial DNA
does. The first results from one
section of a gene on the Y chromosome found no variation in it. Based on the rate that this same
section appears to vary in other primates, the researchers estimated that the
male ancestor common to all modern humans existed around 270,000 years
ago. An extremely low diversity in
other sections of the male Y chromosome has also been reported by other
researchers.61 These results agree well with the
mitochondrial dates and provide independent confirmations of the recent pinch,
bottleneck or replacement model.62 63
After
twenty years of research, Svante Pääbo and others isolated mtDNA from
poorly-fossilized Neandertal remains and reported that there are sufficient
differences between the recovered Neandertal and modern human sequences to
conclude that no Neandertal genetic information survives in modern populations.64 This result lends strong support to a
replacement model, for if extensive intermixing of the established Neandertal
and incoming modern human population had taken place, some genetic evidence of
this should persist in modern mtDNA.
DISCUSSION
Up
to this point we have only reviewed data from the natural sciences that may
bear on the origin of man. Many
Christians, of course, recognize that special revelation is another source of
knowledge that bears on this issue.
We will now turn our attention briefly to the biblical material and
highlight the tension that sometimes occurs when scientific data and revelation
both speak to the same question.
Within
the conservative Christian community there is a wide range of opinion on how to
interpret the biblical material which relates to Creation. Some treat the Creation narrative as if
it stopped after Genesis 1:1 and distance themselves from the host of other
details provided in the next two chapters. This group views the order of creation, special creation of
Adam, Garden of Eden, and Fall as God's allegorical attempts to explain to the
ancient, primitive Hebrews their place in the universe and the origin of sin.65 Others propose a literal
interpretation, taking a simple reading of the English text as historical and
scientifically accurate.66
To
the author it appears that the truth lies somewhere between these
positions. One treads dangerous
territory if one ignores the linguistic and cultural difficulties of attempting
to decipher an ancient text, or if one dismisses any attempt to understand the
passage as historical by inferring that God was so bound by his early audience
that he could convey nothing in Genesis of future relevance to modern man. It seems wisest to propose that this
simple yet sophisticated narrative contains a highly abbreviated summary of the
origin of the Earth and mankind.67 Like other passages, early Genesis does not address a host
of items of interest to contemporary specialists in a particular field: The Bible focuses on salvation history
and only fills in cultural and background details as they are relevant to the
immediate context, not for the curiosity of readers thousands of years later.68 Nevertheless, the events that are described should be understood as real and
historical, especially in light of their implications for mankind. For example, the institution of
monogamous marriage, the origin of sin and the resultant corruption and
mortality of mankind, are all grounded in God’s interaction with a single
male-female pair of human beings, from whom all mankind are descended. If anthropology forces us to replace
Adam and Eve with a global homo erectus population that collectively evolved into modern man, then the
theological foundation for the nuclear family, sin and death appears to be
eroded. The credibility of the
Bible when it speaks on these issues seems to be damaged: If it does not correctly explain the
origin of a problem, why should one trust its solutions?69
As
an example of this integrative tension, let us focus on one specific question,
Was Adam created directly from non-living matter or did he have hominid
parents?
Genesis
2:7 reads, “Then the LORD God formed man of dust from the ground, and
breathed into his nostrils the breath of life; and man became a living
being.” [NASB] In contrast
to previous creative acts where God said, “Let the earth bring
forth....”, the picture here is that God is actively involved in giving
life to a substance that was not previously living.70
It
is striking that most of the specific features in this verse clash with the
anthropological and biochemical models which we have reviewed. Modern scientific theories propose that
man and apes share a common ancestor in the past, that the transition to modern
man took place gradually over thousands of years, and that small groups or
clans of hominids (not one individual) were involved in the process. Some claim that a sudden jump from
non-life to modern man, or even from an individual primitive ancestor to a
modern man, is absolutely ruled out by the diversity seen in the biochemical
and fossil data.
Some
Christian scholars dismiss or excuse these apparently erroneous statements by
asserting that the Bible here is limited by its primitive cultural
setting: The ancient writers and
readers of Genesis were restricted by ancient near eastern cosmology in the way
they viewed themselves and the world.
This mind set so permeates biblical passages like this that no modern
scientific conclusions can be or should be drawn from it. However, conservative scholars hasten
to add that these limitations do not hamper our ability to deduce theological
truths from the text.
This
argument overlooks the fact that ancient cosmologies exhibit considerable
variety. In fact, some creation
stories from neighboring cultures agree better with the modern scientific one
because they lack the “embarrassing” features that a single man was
created and that he was formed from purely non-living matter. The Atra-hasis epic describes the gods
making men from the flesh and blood of a god mixed with clay;71 thus man in this scheme is partially
made from already-living material.
Moreover, the text notes that seven male/female couples were created at
the same time, rather than a single man.72
Neither
can we assume that apes or similar primates were unknown to the primitive
Hebrews. An alabaster statue of an
ape found in Susa [Persia] dates back to the Proto-Urban period (c. 3300 BC).73 Apes (or possibly monkeys) are
mentioned as curiosities in Solomon’s court (1 Kings 10:22) but were
common in Egypt.
Perhaps
the notion of common ancestry between man and other animals was too difficult
or repulsive for the primitive mind (although it is hard to imagine how this
would be more negative than being called “dirt” by God in Gen.
3:19). If the writer of Genesis
had wanted to avoid unpopular concepts, he would not have stressed that all
mankind have a common ancestor! A
multi-Adam model would find easier acceptance because it can justify the
intercultural and interracial hatred that characterizes so much of human
history.
In
light of the comparative literature and cultural knowledge available to us, we
cannot presume that it was impossible for the writer of Genesis to explain to
his ancient audience that God empowered a small group of already-living beings
that shared some common features with apes to develop into mankind. God could have told the story of a
hominid-origin for mankind in a manner the early Hebrews could have
grasped. The fact that he did not
must have some significance.
Returning
to the natural sciences, we find that the available data used to support the
theory of hominid evolution do not show a smooth, gradual transition from
earliest australopithecines to university professor. Two recent, abrupt changes occur in the data: A cultural one
at around 40,000 BC, when art appears and the complexity and variety of
artifacts greatly increases; and an anatomical one at around 130,000 BC when
anatomically modern human remains first appear.74 75 At present either of these transitions
seems sharp enough that we can propose that the special creation of man
occurred in one of these gaps and that it was not bridged by purely natural
means.
One
challenge that the scientific data pose for this recent special creation model
is the clear evidence that a variety of fairly advanced hominids existed on the
earth either immediately before, or contemporary with, modern man. The Bible does not mention any hominids
in the animal population, so if we place Adam at a late gap in the hominid
record (as we have done with either date above) then we must assume that Adam
had non-human contemporaries who made stone tools and used fire. However, as already noted above, DNA
and mtDNA studies strongly imply that these preadamic hominids were replaced by
modern human populations and that little or no interbreeding took place.76 We could speculate that this
preadamic hominid population formed a cultural “safety net” for
Adam and Eve, so that after they were expelled from the Garden they could
survive in the wild by adapting the skills of the hominids around them. These hominids might be the reason for
Cain’s fear that “whoever finds me will kill me” when he is
cursed by God after killing Abel (Genesis 4:14).
The
above consideration of the special creation of man is often ridiculed as an ad
hoc “god of the gaps”
special pleading by secular naturalists and some Christians.77 However, all parties agree that several
gaps and apparently rapid transitions exist in our present-day knowledge of
human prehistory. The problem is
how to deal with them. The special
creationist presumes that at least one gap is real – corresponding to
God’s direct intervention in a world that he normally controls through
what we call natural processes.
The naturalist (either theist or atheist) presumes that all gaps are
only apparent ones because the appropriate fossils have not been found or did
not survive. Consequently he
bridges all gaps with natural processes and sees no need for God to tinker with
his creation (or he sees no need for God at all). This approach requires a strong faith in “naturalism
of the gaps,” but the naturalist argues that this is justified by the
theory of common ancestry.
The
theory of common ancestry is the cornerstone of modern evolutionary
theory. In its simplest form, it
merely states that everything that lived had parents. Once the first living cell formed, all other life on earth
descended from it. Evidence
typically given as proof for this theory is the uniformity of the genetic code
and the high degree of similarity in the biochemical composition, structure and
function of proteins, cells, organs, and higher animals. This similarity can be explained simply
and elegantly by the purely naturalistic mechanism of common ancestry. Everything looks similar because they
share common parents sometime in the past.
Many
scientists view the theory of common ancestry as so obviously true (“the
fact of evolution”) that details about the exact mechanism of
macroevolution (gradual change, punctuated equilibrium, or some yet unknown
process) do not disturb them.78 Such a deep faith in this solely naturalistic model blinds
many to any other possible explanations for this similarity, such as the theory
of a common designer or engineer.
In
the matter of human origins, creationist theories (whether young- or old-earth)
propose that at least one of the observed gaps in the fossil and/or cultural
record is real and is not an artifact of our limited knowledge of the
past. At some point in natural
history God made man by a direct means.
Of course, we can still ask why God made man so similar to modern apes
-- or more importantly -- to the
other hominids that were contemporary with early man.
Perhaps
if we reword this question we can see better how to answer it. Let us instead ask, Does man have to
be different to be proof that God
made him directly? (For the sake
of argument we will ignore the clear differences in language ability and
culture that do distinguish man from animals and other hominids, since a
skeptic would dismiss these as secondary traits). The answer to this is simple: No. We know that many of Jesus’ miracles produced results
that looked like they had a history and were completely natural (e.g., the wine
at the wedding in Cana, the bread used for feeding the 5,000), so some of
God’s miraculous actions can be difficult to distinguish from his
providential ones unless we were there at the time the miracle took place or
have additional knowledge about the circumstances surrounding it. The author views the special creation
of man to be such a case. The
Bible tells us something about the history of man that we may not be able to
deduce from the surviving evidence: Man was a special creation. Why did God choose not to modify an
existing hominid, but make something very similar from scratch? Perhaps to indicate man’s
relative importance before God or to signify that man is the capstone of his
creation. Certainly from reading
Genesis we do not get the sense that there is anything “bad” in
what is created up to this point that God would have an aversion to using as a template
for his final work.
A
rough analogy might be a business letter, dictated to and typed up by a
secretary, but signed personally by the president of the firm. All of the content of the letter comes
from the president, but the signature involves his own personal action and
serves as a stamp of approval, authenticating the contents. Man is God’s personal
“signature” – his own handiwork – on the rest of
creation.
Another
analogy, this time from the ancient near east, involves the building or
refurbishing of a temple. A key
element in this massive undertaking involved the making of the first brick for
the new construction. In an
elaborate ceremony, the king personally mixed the water and mud and molded it.79 Once this first brick was made, then
the manufacture of the rest of the bricks by ordinary workmen could begin. It was not considered demeaning for the
king to stoop to make a brick himself, but was a great honor. The king’s brick did not have a
composition any different than the others; yet because it was made first by the
king personally, it was clearly the most important one. In a similar way, God’s creation
of man is significant not because of man’s unique physical composition,
but due to God’s direct involvement in the process.
While
the theory of a common designer is assumed when studying artwork in order to
determine the style that is characteristic of a given painter, with
self-replicating systems it may be hard to distinguish self-modification from
external adjustment. In cases
where a substantial increase in complexity occurs, external adjustments can
only be ruled out if the mechanism of self-modification is well understood and
if sufficient intermediate forms existed to demonstrate that no discontinuities
occurred. Otherwise it is unwise
to presume that a natural mechanism is the only possible means to bridge the
gap.
If
the goal of science is to determine what actually happened rather than to
construct the most plausible naturalistic rationalization of what could have
happened, then we must apply our presumptions cautiously. As Einstein noted in another context
(quoted in our foreword), the presuppositions in a theory can mask data so that
people no longer see other possibilities.
If “science” is defined so that the actions of a
supernatural agent are considered out-of-bounds or even irrational, we should
not be surprised if it cannot see God’s handiwork through its
self-imposed blindness.
Is
there an objective means to resolve the clash between naturalistic and
interventionist presuppositions?80 More data will certainly fill in the existing gaps in
our knowledge and help us determine which apparent transition is real. Research on several fronts will doubtless
be helpful: More intermediate
hominid fossils will help trace where gradual change in skeletal features did
or did not occur; Continued
comparative studies between human and primate DNA will give us a more detailed
picture of our similarity and provide more clues as to its significance; Archaeological discoveries in biblical
studies may find new material that will help us better understand early
Genesis.
For
the present the author feels no compulsion to bridge all of the gaps in hominid
evolution with naturalism instead of theism. The clear break from hominid to human culture, the high similarity of modern human DNA that
implies even to secular minds a recent “pinch” in population, and
the overall fragmentary nature of our present knowledge, should make us
hesitant to ascribe the works of a common designer to common parents.
In
conclusion, it is worth noting that both science and theology benefit from the
tensions that arise when the interpretations of the data from each field
conflict. Neither field can claim
to be an absolute guide to truth apart from the other: Theology learned from
its geocentric/heliocentric debates with the natural sciences to be cautious in
its interpretations, because some biblical language is phenomenological, just
as many idioms of regular speech are.81 Likewise science learned to be cautious in its metaphysical
presuppositions when it became clear that the universe is not eternal but
appears to be created, something Christian theologians have argued for hundreds
of years.82 In the case
of human origins, no obvious resolution of the tension between the models seems
possible at present. By watching
for blind spots in our presuppositions, double-checking the validity of our
data and seeking more of it, perhaps we can arrive at a consensus that is more
charitable to all.
[2]
Because it touches
closest to home, the controversy surrounding the creation / evolution debate
becomes noisiest when the question of human origins is broached. Certainly
if macroevolution only concerned itself with bacteria, fish and rodents,
its discussion would take place in the footnotes of biology journals; but
since it ventures to offer a scientific opinion about who we are and what
was involved in the process of getting us here, it commands the attention
of society at large.
This occurs because the meaning
and value that we assign to life are linked to our models for its origin: For
example, some argue that we evolved by a meaningless collision of molecules
and have no more significance than insects who hatch, eat, copulate and
die; while others hold that we are a unique special creation formed to
glorify God. Because we connect our significance with
our pedigree, the following review of the physical data on human origins
will include some consideration of its metaphysical fallout.
[3]
Thus
this survey is offered in the hopes that it will not (further) divide
the conservative Christian community, but that it will illuminate the
common ground where all can agree and respond to the far more dangerous
naturalistic assumptions inherent in most secular models. Of
course, the most profound of these is the bald assertion that the created
order lacks meaning or purpose. When
we look at the complexity of the final products, no matter how we model
the mechanism of their initial formation, how can we permit
anyone to deny there is design to the universe? Arguing for the existence of a Designer,
and hence for a purpose to his work no matter what mechanism he uses
to carry out his will, should be our priority. [Tie back into the introduction.....]
End
notes: