
The Functions of Introns:
From Junk DNA to Designed DNA
Jerry Bergman*
Northwest State College
jdbrg@bright.net
22-600 State Rt. 34
Archbold, OH 43502-9542
The research on functions for introns in the cell is reviewed. Darwinists once generally argued that nonprotein coding DNA are relics of once-functioning genes or useless "junk" DNA that strongly argued against design of the genome. The fact that an enormous quantity of cell resources and energy is invested in these putative vestigial structures, especially in the complex intron splicing mechanism, argues that introns have important biological functions including a means of facilitating genetic diversification. Evidence now exists that introns have many functions, including for regulation and structural purposes, and that many of the roles now hypothesized for introns are plausible but need further elucidation. The author concluded that the new knowledge related to introns supports the intelligent design world view.
The past decade has witnessed an explosion of discoveries in the area of molecular biology that has numerous practical and major implications for the creation-evolution issue. A critical requirement for evolution is the existence of large sections of extra nonfunctional DNA that can mutate to produce potential genes upon which natural selection can act. Bacteria usually contain only a few thousand genes (E. coli has 4,253), and humans have about 35,000. Consequently, evolution from protobacteria to humans requires a mechanism that can add functional genes.
One currently popular hypothesis to account for this increase in the genome size is that they are the remnants of the original development of genes, or that many DNA bases were added during evolution through such sources as viruses or inappropriate DNA duplication.1 Called evolution's fuel, the theory argues that extra DNA allowed natural selection to gradually convert some of the nonfunctional DNA into functional genes.2 The redundancy theory requires the organism to possess extra DNA in addition to its functional genes to allow it to evolve new genes that can produce new protein and cell structures. The redundancy theory suggests that most genes were once noncoding DNA and/or nonsense genes that produced nonfunctional protein which was either useless and rapidly recycled or caused harm to the cell.
Production of nonfunctional protein would result in a tremendous waste of cell energy and materials both in manufacturing the useless proteins produced by evolving genes and as a result of their being cut up and recycled by the cell's proteolytic system. Proponents of the redundancy theory speculate that some of these nonsense genes eventually produced a protein that conferred a survival advantage on the organism that possessed it. Last, natural selection fine-tuned the DNA and caused the protein it produced to become increasingly useful to the organisms' survival. For this process to occur, a large number of nonfunctional coding genes must have existed. Furthermore, without sufficient genetic raw material, evolution could not occur by this process. Much raw material must have existed and evolved into coding genes that produced a phenotype which conferred a selection advantage to the organism.3 This belief is supported by the belief that an estimated 95% or more of eukaryote DNA has either nonprotein-coding functions or no known function at all.
The DNA sections that have no known function are used as evidence for this theory. Nonfunctional DNA not only provides the raw material required for evolution, but it supports the Darwinist's view that the genome shows no evidence of design. Evolutionists reason that an intelligent designer would not produce a genome containing long sections of useless DNA that waste enormous amounts of materials and energy by duplicating huge sections of nonfunctioning DNA during each cell division that build useless (or even harmful) proteins. Sagan and Druyan state: "Some, maybe even most, of the genetic instructions must be redundancies, stutters, untranscribable nonsense ... [which prove that] deep imperfections [lie] at the heart of life."4 This view also supports the position that evolution is a blind, purposeless process except if it facilitates an organism's ability to pass on its genes.
This view must be evaluated in light of the fact that the history of science is replete with now discarded theories that once supported Darwinism but increasing knowledge has rendered obsolete. Examples include vestigial organs (the claim that in humans 180 organs and structures were evolutionary leftover's such as the pineal and the thymus), atavisms (the theory that certain conditions such as trisomy 21 were evolutionary throwbacks--trisomy 21 persons were called Mongolian idiots for this reason), and the biogenetic law (the belief that the embryo- logical development of an organism passes through its evolutionary history). Likewise new research is beginning to overturn the view that most of the genome has no function. This is reflected in the following comment on the Sagan and Druyan statement cited above:
Such comments are commonplace in the biological literature--although perhaps less common than they were a few years ago. The reason? Geneticists are discovering functions for what used to be apparent genetic debris.5
In contrast to the requirement of evolution for large amounts of useless DNA, the research explosion on DNA has revealed evidence that much of this DNA is necessary, or even critical, for life. Although many of these studies are preliminary, and many other uses for noncoding DNA likely will be found in addition to those noted here, most recent discoveries support the design world view. Recent research also supports the view that, though over-designed (viz., possessing more information than is necessary for survival) the entire genome is or once was functional and exists for a biological purpose--a conclusion that supports the intelligent design hypothesis.
Introns
The two major divisions of DNA that are transcribed into RNA are protein-coding sections called exons, and nonprotein-coding sections called introns. Introns consist of large stretches of DNA whose biological functions are only beginning to be elucidated. All genes begin with exons (the protein-coding segments), but most have a variable number of introns within them that alternate with the exons. Introns were discovered in 1977 as a result of observing that the mRNA used to code for proteins was almost always shorter than the DNA from which it had been transcribed.6
The mRNA was eventually found to be shorter because it lacked the noncoding sequences (introns) between the coding regions (the exons) on the DNA.7 It was discovered that introns were normally removed by splicing enzymes before mRNA (messenger RNA), rRNA (ribosomal RNA) and tRNA (transfer RNA) can complete their functions in the cell. Because introns interrupt the nucleotide sequences, they first were called interrupted genes. The "int" in intron refers to intervening because introns always exist between exons. In eukaryotes, intron removal and splicing is completed within the nucleus. Sequences that code for protein are called exons because they travel (exit) outside the nucleus to code for proteins, and thus are the DNA sequences that are expressed (the prefix ex in the term exon is from expressed).
The Process of Removing Introns Shows Evidence of Design
When mRNA is transcribed
from DNA (Figure 1), both the exon and intron noncoding sequences are
transcribed into a macromolecule known as a heterogeneous nuclear RNA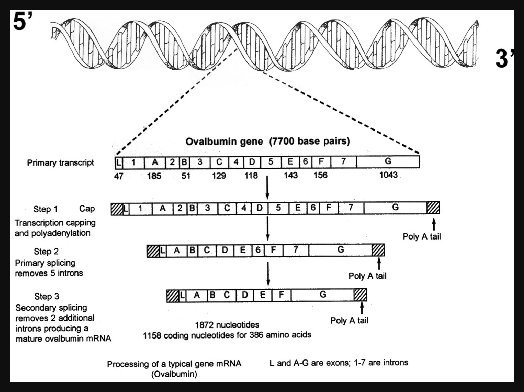 (hnRNA), or immature RNA. Before the RNA exits the nucleus, the introns must be
removed by a precise cleavage-ligation reaction called splicing, thereby
producing a functional mRNA.8 Enzymes and small
ribonucleoprotein structures exist in all eukaryotic cells to assist in removing
introns. In most cases, type I introns are removed in the nucleus by a complex
splicing machine composed of small nuclear ribonuclear proteins (SNRP).
SNRPs consist of 60-300 small nuclear RNA (snRNA) nucleotides in an
"intimate alliance with a bouquet of proteins."9
(hnRNA), or immature RNA. Before the RNA exits the nucleus, the introns must be
removed by a precise cleavage-ligation reaction called splicing, thereby
producing a functional mRNA.8 Enzymes and small
ribonucleoprotein structures exist in all eukaryotic cells to assist in removing
introns. In most cases, type I introns are removed in the nucleus by a complex
splicing machine composed of small nuclear ribonuclear proteins (SNRP).
SNRPs consist of 60-300 small nuclear RNA (snRNA) nucleotides in an
"intimate alliance with a bouquet of proteins."9
The process of intron removal involves a precise looping process controlled by a specific nucleotide sequence that abuts the exons.10 Almost all known introns are identified by specific consensus sequences [GT at the start or donor (3') end and AG at the other end, called the acceptor (5') site] which help to identify introns for removal. The term "consensus sequence" is employed because, although the sites may consist of more than just GT donor and AG acceptor bases, so far as is known these sequences are common to all eukaryotic organisms.11 Consensus sequences are DNA segments that use similar base sequences in different genes within a single gene family or across many different species for one specific function.
After the introns are cut out, the exon RNA is spliced back together by RNA ligase so that the final mRNA used to code polypeptides normally consists only of exons. In Type II introns, RNA itself functions as an enzyme called a riboenzyme or ribozyme. These enzymes require a divalent cation (usually magnesium) to function. The process of removing introns involves a complicated reaction pathway that includes RNA intermediates called lariats, branch connecting points, and a large number of accessory proteins.12
Number and Length of Introns
The sections of the human genome sequenced thus far provide an estimate of the genome's composition. It appears that over 90% of the DNA bases consist of repetitive noncoding regions, introns, and regions between genes that have no known function.13
The number of introns in transcribed RNA ranges from zero in histone genes to an estimated 2.5 million bases in the protein dystrophin.14 The dystrophin gene has over 75 introns and only 0.5 per cent (11,000 bps) of the gene consists of exons.15 A defective dystrophin gene is the major cause of Duchene Muscular Dystrophy. Another example is approximately 98% of the gene that causes cystic fibrosis consists of introns.16 The large number of introns in the genome is revealed by evidence that indicates the majority of base pairs in most genes consists of introns. The length of one intron varies from 31 nucleotides in an SV-40 gene to over 210,000 in the human dystrophin gene,17 although 65 to 20,000 nucleotides is more typical.18
Roles of Introns in the Cell
Indications that introns have a use include the observation that "the cell puts a huge amount of its energy into the creation of these introns, then discards them ... Nature would not go to all that trouble without a reason."19 Why "most genes of higher organisms are broken up into short exons separated by huge stretches of seemingly useless DNA" is particularly puzzling because "the introns are all painfully transcribed into RNA only to be snipped out by snRNPs and thrown away almost immediately."20 Evolutionist Patrusky says: "Nature, for reasons as yet unknown, created the intron, and evolution has chosen to keep it ... and ultimately has found new ways to use it."21
An important finding which may help researchers to understand the role of introns is the more primitive and simple the organism generally the fewer the introns. Although the majority of plant and animal genomes have not yet been examined, introns have been found in the vast majority of eukaryotic genes and likely exist in all eukaryotes.22 Wills speculated that fewer introns exist in prokaryotic organisms because bacteria need streamlined genomes, or may have less need for whatever functions introns serve. He also speculated that the putative early organisms may have lost their introns through evolutionary development.
The fact that all higher organisms possess introns, and the more complex organisms possess a higher proportion of introns, indicates that they serve at least minor, and possibly major, functions. Wills says: "Carrying this enormous extra baggage of unnecessary DNA each generation seems a heavy price to pay for the privilege of the occasional bout of exon shuffling."23 Many reasons now exist to conclude that introns are not found in cells only because they are "intrinsically capable of replication and therefore can stick around" as once believed.24 Furthermore, research has found a high level of conservation in some introns, indicating that they have some selective advantage, that is a useful function.25 How many introns display this conservation is not yet known.
This raises the question, "If introns produce a major selection advantage and consequently are characteristic of higher, more developed organisms, what could explain their loss in lower organisms?" Variety is critical for species survival, and producing variety is especially difficult in animals with small genomes, the very creatures which must have possessed the variety required by evolution in order to evolve. Putative ancient bacteria evidently would possess little capacity for variety if the absence of introns (and other structures) in modern bacteria was also the situation in early bacteria.
The former belief that introns were useless was once so prevalent that many felt it would be a major waste of resources for the human genome project to sequence these long stretches of "meaningless DNA." The human genome project involves sequencing the estimated 35,000 genes and over three billion base pairs at a cost of over $3 billion.26 The National Science Foundation priority is now on sequencing exons, but eventually the entire code will likely be determined partly because of accumulating evidence that the entire genome has a function. A problem in genetic research is that genes in which introns are left in may be so long and unwieldy that they cannot be carried by a vector virus or plasmid. When the significance of introns is understood better, this problem can be dealt with more effectively.27
The New View of Introns
It is now recognized that introns are "a complex mix of different DNA, much of which are vital to the life of the cell."28 As their functions are being determined, the relationship of introns to cancer and their use as tumor markers is also being explored. Several functions for introns have already been identified, and evidence for a role for them is indicated by the finding that some intron alterations are directly related to the development of cancer.
Early insight in intron function was a result of the finding that many noncoding DNA sequences are not random base pairs, but have certain features in common with human language.29 The finding that introns manifest the same complex patterns of communication found in human speech supports the supposition that they carry functional information. Of course, the existence of a pattern does not in itself provide direct evidence for a function, but it does indicate a potential systematic cause exists which produces the pattern found.
Alternative Splicing
The problem of identifying introns was compounded by the discovery that alternative readings of the genetic code exist in which "introns" function as exons.30 Some DNA behave as exons when expressed by one pathway, but as introns when expressed by another pathway.31 Both pathways can operate simultaneously, resulting in greater protein product variety.32
Termination codons are also sometimes deliberately bypassed, allowing the coding of a part of an intron in order to produce a specialized protein. Variations in intron removal splicing patterns can be used to create two or more distinct mRNAs which code for different polypeptides. This allows one mRNA code to be used to produce a variety of polypeptides.33 Splicing variations are controlled by regulator proteins designated as SR proteins. SR proteins determine which splicing pattern predominates in a specific cell at any given time.34
Another example of an alternate reading system that can make use of introns is frame shifting, which causes the normal grouping of nucleotides to be altered so that the DNA is read in a completely different way. A third type of alteration which may involve introns is base hopping in which large DNA segments are skipped when the mRNA is produced. These observations reveal that genes are far more complex than our earlier understanding of the genetic code first indicated.35
Some introns also function as control sequences in the process of chromosome X inactivation which is necessary for dosage compensation in females. The Sxl gene is the master regulator in the sex determination pathway of some organisms and functions by regulating how the introns are cut out of mRNA. It produces a functional protein only in females that culminates in the development of female structures.
The Sxl gene is also critical for dosage compensation by blocking mle and msl genes, which make a protein that increases certain X chromosome proteins.36 Sxl functions by blocking removal of the introns in females, preventing females from producing functional msl-2 protein. The msl-2 gene is also controlled by removal of an intron in males, but not in females.37 Males lack an Sxl gene, and therefore process the msl-2 transcript properly:
The fruit fly provides an interesting example of two- way mRNA splicing. In this animal, the differences between males and females are controlled by two sets of regulatory proteins, one set specific to the male and one to the female. The two sets of proteins are coded by the same set of genes; the sex differences are largely due to different patterns of RNA splicing.38
Nested Genes
Some introns have been found to be separate genes located within the reading frame of introns and are often transcribed from the opposite direction as the introns. Many examples have been found, and likely many more exist. The neurofibromatosis type I gene contains three nested genes, all transcribed in the direction opposite that of the gene.39 These three nested genes appear to have no functional relationship to the neurofibromatosis gene. An intron in the blood clotting factor VIII gene in the human X chromosome contains an embedded gene that is also transcribed in the opposite direction, and a second embedded gene that is transcribed in the same direction as factor VIII gene.40
The discovery that some DNA function as exons when expressed by one pathway, but as introns when expressed by another, blurs the distinction between introns and exons. The finding that nucleotides cut out introns in one splicing pattern which become part of the functional mRNA as exons in another splicing pattern also indicates that a precise method to differentiate introns from exons exists.
Another possible example of a control function for introns was inadvertently discovered from research on transgenes developed for gene therapy. In endeavoring to replace a malfunctioning gene with a normal allele, a research team headed by Oliver Smithies of the University of Wisconsin removed the introns after they had determined that the genes were not expressed. They found that "the shortened versions of the HPRT gene commonly being used in such experiments worked very badly in cells recently taken from mouse embryos, although they did work well in cells taken from mice years before and grown in culture."41 Conversely, when the researchers left in the introns, they found that the genes worked properly in "fresh" cells. This research indicates that at least some introns may have an important role in controlling the expression of exons.42 Whether this putative regulation system affects tumorigenesis is unknown.
Although introns are routinely edited out, Ron James and his team at Pharmaceutical Proteins Ltd. injected mouse eggs with an alpha-1 antitrypsin gene that included some of the introns in transgenic sheep. James states: "We left some of these random bits of DNA in the gene, essentially as God provided it," and as a result "high yields resulted."43 In this case, the introns enhanced the production of protein, resulting in a breakthrough crop of transgenic sheep. This research also supports a facilitator role for introns.
Many diseases are caused by mutations in the noncoding portion of the gene, such as in the flanking or start-stop sequences. Examples include hemoglobin diseases caused by damage to a noncoding section of DNA that contains sequences identical to the normal hemoglobin exons. The hemoglobin protein produced by the gene evidently will not function if a mutation occurs in certain non-exon DNA.44 Other noncoding portions also have an impact on the organism's function, and it is possible that the introns have a role in reducing or correcting these mutations.
Regulatory Role of Introns and Cancer
At least a dozen studies shave found evidence that introns are either directly or indirectly involved in cancer causation. Examples include evidence that introns are involved in transcriptional regulation of apoprotein B, E, and A-II45 and that introns may be involved in regulating neoplasm development.46
Cytolytic T lymphocyte clones used to study melanomas found the gene coding for the antigen recognized by the cytolytic T lymphocyte was the same gene which codes for N-acetylglucosaminyl- transferase V. The antigenic peptide recognized by the cytolytic T lymphocyte was found to be encoded by a sequence located inside an intron. The researchers found that the mRNA containing the introns coding for the antigen was not found at significant levels in normal tissues but was observed to be present in close to half the melanoma tissues studied. The researchers concluded that a promoter located near the end of the relevant intron was activated in melanoma cells, resulting in the production of an mRNA that codes for the antigen.47
Defective glutathione S-transferase and N-acetyl- transferase enzymes have been associated with an increased risk of developing both lung and bladder cancer. The research results are inconsistent, though, and several studies have failed to find associations. According to some studies, the lung cancer risk is elevated up to 40-fold in subpopulations that contain both the high-risk cytochrome P-450 type A1 and glutathione S-transferase M1 genotypes which are a result of mutations in introns or other silent areas of DNA. One study on the glutathione S-transferase M3 gene found a mutant three-base deletion in intron 6 of the wild type glutathione S-transferase allele.48 This defect may be related to neoplasm development, but exactly how is unknown.
Megonigal, et al. used panhandle PCR to clone MLL genomic breakpoints in two pediatric treatment-related leukemias.49 The panhandle PCR identified a fusion of MLL intron 6 with a previously uncharacterized sequence in MLL intron 1 which the researchers concluded was consistent with a partial duplication. The breakpoints in both cases were located in Alu repeats, suggesting that the Alu sequences were an important contributor to the rearrangements they found.
Malkinson and You have hypothesized that introns of genes whose products influence tumor development can affect cancer incidence and that
a large number of regulatory mechanisms ... control key growth and differentiation steps: ... The ability of each of these host defenses to withstand mutation and a consequent loss of function may be controlled, in part, by their intronic structure. Proto-oncogenes, tumor suppressor genes, cell-cycle regulatory genes, simmune defense systems, and the protease/ antiprotease systems that influence metastasis are all candidates for having polymorphisms within their introns that affect gene behavior.50
Conclusions about introns and cancer are difficult for several reasons, including the fact that it must be determined if mutations in the introns actually contribute to tumorigenesis progression, or if this damage is a result of collateral or unrelated damage. The associations found between introns and cancer may occur by chance, or it may be due to an unknown role of introns or other putative silent areas of DNA. Numerous other studies indicate a regulatory function for introns:
Introns are becoming more widely recognized as having important gene-regulatory roles, such as containing enhancer or silencer elements. While many intronic polymorphisms may be of trivial consequence ... we believe that variation in the number of intronic 37-bp copies in the Kras will serve as an interesting model system for examining how inherited variations in the copy number of oligonucleo- tide regions within an intron affect the etiology of diseases.51
Among the many other intron genetic variations that have been linked to cancer (in this case lung cancer) is a K-ras intron variation involving tandem repeats in the H-ras 3'-untranslated region.52 Mutations in introns may influence neoplasm development because mutations that occur in introns (or in the DNA sequences that flank the gene) can affect the various steps required for normal expression of the gene even though the intron codes may not be detectable by studies of the protein product of the gene.53 If the defective expression of a gene product is a tumor suppressor gene or a proto-oncogene, cancer may result.
Regulatory functions of introns may involve controlling gene activity in different developmental stages or responding to immediate biological needs by controlling local gene expression.54 This function of introns could occur if, as one theory indicates, exons code for a domain, a polypeptide unit that has a discreet function such as binding to a membrane, or to the catalytic site of an enzyme or serving as a structural unit of a protein.
Some evidence also exists that introns structurally stabilize the pre-mRNA to protect it against degradation. Margulis and Sagan noted that DNA is packed into a chromosome so effectively that it is 1/8,000 of its former size after packing.55 They concluded from studies on ciliates that at least some introns, and possibly other noncoding DNA, may be involved in DNA packaging in eukaryotes and perhaps in some prokaryotes.56
Exon Shuffling
Introns also produce a critically important life trait called phenotypical variety by regulating or facilitating the transposition of exons.57 Evidence for this role includes the observation that introns commonly are located at or near the boundaries between different domains in the DNA molecule. Called exon shuffling, this mechanism helps to produce genetic and phenotypical variety in an organized way by producing new exon combinations and thus greater genetic diversification. This would enable the production of a variety of poly- peptides from a single gene.
Bryk and Belfort concluded that ectopic crossovers (nonspecific pairing of DNA) could cause shuffling to occur in an open reading frame so that "double crossovers between separate elements" resulted. They suggested that this process would "extend the range of viable recombinational possibilities because they leave undisturbed the regions between the homologous elements."58 This finding supports the conclusion that some introns act as recombinogens that function to increase the level of genetic exchange and consequently controlled phenotypic variety.
Introns may also contribute to greater genetic diversity due to their role in crossing over, which is one of the "main sources of genetic variety in eukaryotes."59 The role of introns in producing genetic variety includes the lengthening of a DNA strand considerably, thereby increasing the number of points where crossing over can occur between two gene alleles. Introns may also assist in the recombination process to ensure that exons are shuffled in an orderly manner to produce appropriate exon combinations.
Another role for introns in producing variety involves introns related to retroposon elements that are able to move DNA from one part of the genome to another in a way similar to how retroviruses (including HIV, and many cancer viruses) function. Retroviruses carry a gene that produces a reverse transcriptase enzyme that codes DNA from mRNA. Introns accompanied by flanking exon sequences use a highly efficient homing process to move to its intron-less alleles at efficiencies approaching one hundred percent.60 Specific mechanisms that introns use to achieve this still are not understood. Although some introns resemble transposable genetic elements, most do not. The function of introns to produce phenotypic variety could support both an intelligent design interpretation (they are complex designed mechanism that produce limited variations in certain structures) or evolution (they provide variety for natural selection to operate on). Functional theories argue that introns and other noncoding
DNA has a sequence-independent function by virtue of its sheer bulk. Most functional theories assert that cell size is adaptively important (for which there is much evidence;61) and that the genome-size-cell- volume relationship is the key to explaining the continued presence of non-coding DNA. The most specific of the functional theories is the skeletal DNA hypothesis, which states that the mass of DNA (in conjunction with its folding pattern) directly determines the nuclear volume. In order to maintain a balance between the rates of nuclear RNA and cytoplasmic protein synthesis during growth ... cells of differing volumes must have a constant cytonuclear ratio.62
This ratio tends to be optimized in all cells because of certain cellular requirements. Cavalier-Smith have found that their functional theory applies to both unicells and multicells, but its application to the latter is more complex due to the extra complications in multicells and protists with multiple nuclei or multiple fission.63
Conclusions
Various known and possible functions of noncoding DNA were briefly reviewed with special emphasis on introns. The research reviewed in this paper has caused a number of investigators to conclude that the hypothesis, that large amounts of DNA are nonfunctional, may be erroneous. As research continues to elucidate the structure of the genome, it seems that most DNA will be found to have a function, although some DNA no doubt will prove to be more important than other DNA. Furthermore, some useless DNA which has little effect on survival may be a result of corruption of genes due to entropy. Some useless DNA may also be the remains of virus or transposon DNA inserted into the chromosomes in the past.64
No clear evidence yet exists that introns support an evolutionary origin for the genome. It appears that problems develop with noncoding DNA only when mutations or other damage to the system occurs. We now know that DNA sequences not only carry information for making proteins, but also play many supportive roles in protein synthesis including manufacturing transfer RNAs that help to assemble amino acids into proteins and rRNA, as well as other forms of RNA.
In the past, evolutionary geneticists, once uncertain as to what this apparently superfluous DNA does, referred to introns and other noncoding DNA as "junk." Evidence is now being accumulated which indicates that much or most of this DNA may not be junk, but critical for life itself.65 If functions for most or all of the noncoding DNA is found, Darwinism would be without the raw material needed to produce new genes by mutations that can be selected for evolution to occur. Furthermore, much of this new information on the complexity of the genome elegantly provides evidence for both intelligent design and for the concept of irreducible complexity.
g
Notes
Thomas H. Eickbush, "Molecular Biology: Introns Gain Ground," Nature 404 (2000): 940-3. Bob Cory, "Evolution's Fuel," New Scientist (April 22, 2000): 50, 51. Richard Dawkins, Unweaving the Rainbow: Science, Delusion and the Appetite for Wonder (Boston: Houghton Mifflin, 1998). Carl Sagan and Ann Druyan, Shadows of Forgotten Ancestors: A Search for Who We Are (New York: Random House, 1992), 128.5Paul Nelson, "The Junk Dealer Ain't Selling that No More," Origins & Design
18, no. 2 (1997): 21. J. D. Watson, M. Gilman, J. Witkowski, and M. Zoller, Recombinant DNA (New York: W. A. Freeman and Company, 1992). W. S. Klug and M. R. Cummings, Concepts of Genetics (Columbus: Charles E. Merrill Publishing Company, 1983), 81.8E. A. Birge, Bacterial and Bacteriophage Genetics: An Introduction (New York: Springer-Verlag, 1981), 14.
B. Patrusky, "The Intron Story," Mosaic 23, no. 3 (1992): 22-43; and Peck Ritter, Biochemistry: A Foundation (New York: Brooks/Cole Pub., 1996), 710.10S. R. Downie, R. G. Olmstead, and G. Zurawski, "Six Independent Losses of the Chloroplast DNA rp12 Intron in Dicotyledons; Molecular and Phylogenetic Implications," Evolution
45, no. 5 (1991): 1245-59.11Lynn Jorde, John Carey, and Raymond White, Medical Genetics
(St. Louis: Mosby, 1997). K. P. Williams, D. N. Fujimoto, and T. Inoue, "A Region of Group I Introns that Contains Universally Conserved Residues but Is Not Essential for Self-Splicing," Proceedings of the National Academy of Sciences of the United States 89 (1992): 10400-4. R. Nowak, "Mining Treasures from 'Junk DNA'," Science 263 (1994): 608. T. R. Burglin, T. M. Barnes, M. D. Adams, C. Fields, and J. C. Venter, "Introns in Sequence Tags," Nature 355 (1992): 632. M. Cummings, Human Heredity (St Paul, MN: West Publishing Company, 1994), 973-5. C. Wills, Exons, Introns, and Talking Genes: The Science Behind the Human Genome Project (New York: Basic Books, 1991), 239.17Watson, et al., Recombinant DNA
, 138.18Peck Ritter, Biochemistry: A Foundation
(New York: Brooks/ Cole Pub., 1996), 711.19C. C. Kopezynski and M. A. T. Muskavitch, "Introns Excised from the Delta Primary Transcript Are Localized Near Sites of Delta Transcription," The Journal of Cell Biology
119 (1992): 503. Wills, Exons, Introns, and Talking Genes, 112.21B. Patrusky, "The Intron Story," Mosaic
23, no. 3 (1992): 33.22C. Wills, The Wisdom of the Genes: New Pathways in Evolution
(New York: Basic Books, 1989), 178. Ibid. L. Margulis and D. Sagan, Origins of Sex: Three Billion Years of Genetic Recombinations (New Haven, CT: Yale University Press, 1986), 23.25L. Sun, Y. Li, A. K. McCullough, T. G. Wood, R. S. Loyd, B. Adams, J. R. Gurnon, and J. L. Van Etten, "Intron Conservation in a UV-Specific DNA Repair Gene Encoded by Chlorella Viruses," Journal of Molecular Evolution
50, no. 1 (2000): 82-92; and Joanna Marchant, "Precious Junk: Garbage DNA Has its Uses," New Scientist (February 2000): 16.26Watson, et al., Recombinant DNA
.27Benjamin Lewin, "The Organization of Interrupted Genes
," chap. 35 in Genes IV (New York: Oxford University Press, 1990); and A. Coghlan, "The Cryptographer Who Took a Crack At 'Junk' DNA," New Scientist 1879 (June 26, 1993): 15. Nowak, "Mining Treasures from 'Junk DNA'," 608.29R. N. Mantegna, S. V. Buldyrev, A. L. Goldberger, S. Havlin, C. K. Peng, M. Simons, M. Stanley, and H. E. Stanley, "Linguistic Features of Noncoding DNA Sequences," Physical Review Letters
73 (1994): 3169-72. George Malacinski and David Freifelder, Essentials of Molecular Biology (Boston: Jones and Bartlett Publishers, 1998). Klug and Cummings, Concepts of Genetics, 83. C. M. Asy, B. Williams-Ahlf, J. Naggert, and S. Saith, "Intron-Exon Organization of the Gene for the Multifunctional Animal Fatty Acid Synthase," Proceedings of the National Academy of Sciences of the United States 89 (1992): 1105-8. W. F. Doolittle and A. Stoltzfus, "Genes-in-Pieces Revisited," Nature 361 (1993): 403. Ritter, Biochemistry: A Foundation, 710.35Philip J. Farabaugh, "Alternate Readings of the Genetic Code," Cell
74 (1993): 591-6. Nigel Williams, "How Males and Females Achieve X Equality," Science 269 (Sept. 29, 1995): 1826-7. Ibid., 1827. N. Campbell, L. G. Mitchell, and J. B. Reece, Biology Concepts and Connections (Redwood City, CA: Benjamin Cummings Publishing Co., 1994), 203. Jorde, et al., Medical Genetics, 21. Ibid., 89.41Wills, Exons, Introns, and Talking Genes
, 65. M. Adachi, R. Watanabe-Fukunaga, and S. Nagata, "Aberrant Transcription Caused by the Insertion of an Early Transposable Element in an Intron of the Fas Antigen Gene of 1pr Mice," Proceedings of the National Academy of Science of the United States (1993), 1756-60. J. Levine and D. Suzuki, The Secret of Life: Redesigning the Living World (Boston: WGBH, 1993), 169.44Jorde, et al., Medical Genetics
.45F. L. Chartier, J. P. Bossu, N. Vu-Dac, J. C. Fruchart, and B. Laine, "Involvement of Intronic Sequences in the Transcriptional Regulation of Apolipoprotein B, E and A-II Genes," Zeitschrift
fur Gastroenterologie 34, suppl. 3 (June 1996): 44-5. Y. Guilloux, S. Lucas, V. G. Brichard, A. Van Pel, C. Viret, E. De Plaen, F. Brasseur, B. Lethe, F. Jotereau, and T. Boon, "A Peptide Recognized by Human Cytolytic T Lymphocytes on HLA-A2 Melanomas Is Encoded by an Intron Sequence of the N-acetylglucosaminyltransferase V Gene," Journal of Experimental Medicine 183, no. 3 (Mar. 1, 1996): 1173-83.47Ibid.
48A. Inskip, J. Elexperu-Camiruaga, N. Buxton, P. S. Dias, J. MacIntosh, D. Campbell, P. W. Jones, L. Yengi, J. A. Talbot, R. C. Strange, et al., "Identification of Polymorphism at the Glutathione S-transferase, GSTM3 locus: Evidence for Linkage with GSTM1*A," Biochemical Journal
312, pt. 3 (December 15, 1995): 713-6. M. Megonigal, E. Rappaport, D. Jones, C. Kim, P. Norwell, B. Lange, and C. Felix, "Panhandle PCR Strategy to Amplify MLL Genomic Breakpoints in Treatment-Related Leukemias," Proceedings of the National Academy of Sciences 94, no. 21 (1997): 11583-8.50Alvin Malkinson and Ming You, "The Intronic Structure of Cancer Related Genes Regulates Susceptibility to Cancer," Molecular Carcinogenesis
10 (1994): 64. Ibid.52G. Manenti, L. DeGregorio, S. Pilotti, et al., "Association of Chromosome 12p Genetic Polymorphisms with Lung Adenocarcinoma Risk and Prognosis," Carcinogenesis
18 (1997): 1917-20. H. Sugimura, N. E. Caporaso, R. V. Modali, et al., "Association of Rare Alleles of the Harvey Ras Protooncogene with Lung Cancer," Cancer Research 50 (1990): 1857.54J. Abelson, "Recognition of tRNA Precursors: A Role for the Intron," Science
255 (1992): 83.55Margulis and Sagan, Origins of Sex,
115.56M. I. Baldi, E. Mattaccia, E. Bufardeci, S. Fabbri, and G. P. Tocchini-Valenti, "Participation of the Intron in the Reaction Catalyzed by the Xenopus tRNA Splicing Endonuclease," Science
255 (1992): 1404-8.57Wills, Exons, Introns, and Talking Genes
, 172. M. Bryk and M. Belfort, "Spontaneous Shuffling of Domains Between Introns of Phage T4," Nature 346 (1990): 396. Campbell, et al., Biology Concepts and Connections, 203. M. Belfort, "An Expanding Universe of Introns," Science 262 (1993): 1009. T. Cavalier-Smith, "Cell Volume and the Evolution of Eukaryote Gene Size," in The Evolution of Gene Size (Chichester, UK: Wiley, 1985). M. J. Beaton, and T. Cavalier-Smith, "Eukaryotic Non- Coding DNA is Functional: Evidence from the Differential Scaling of Cryptomonad Genomes," The Royal Society 266 (1999): 2053-4. Cavalier-Smith, "Cell Volume and the Evolution of Eukaryote Gene Size," 105-84; and ------, "Coevolution of Vertebrate Genome, Cell, and Nuclear Sizes," in Symposium on the Evolution of Terrestrial Vertebrates (Modena, Italy: Mucchi, 1991), 51-86. David Clark and Lonnie Russell, Molecular Biology (Vienna, IL: Cache River Press, 1997), 327. A. F. Smit, "The Origin of Interspaced Repeats in the Human Genome," Current Opinion in Genetics & Development 6, no. 6 (1996): 743-8.s; G. Berx, F. Nollet, K. Strumane, and F. van Roy, "An Efficient and Reliable Multiplex PCR-SSCP Mutation Analysis Test Applied to the Human E-cadherin Gene," Human Mutation 9, no. 6 (1997): 567-74; M. S. Clark and W. J. Wall, Chromosomes (London: Chapman and Hall, 1996); and K. F. Schmidt, "A DNA Structure that Tags Genetic Junk?" Science News 144 (1993): 21.