
The
Trilobite: Enigma of Complexity
A Case for Intelligent Design
Arthur V. Chadwick*
1215 Honeysuckle
chadwick@swau.edu
Keene, TX 76059
Robert F. DeHaan*
7714 McCallum St.
RDehaan237@aol.com
Philadelphia, PA 19118
Perspectives on Science and Christian Faith 52.4 (December 2000): 233-241This paper will demonstrate that we can know in detail the molecular biology of one of the earliest forms of metazoa, the Trilobite. We will reconstruct its molecular biology in order to show the existence of all of the major innovations found in the spectrum of life on the earth today. In the absence of evidence for the evolution of complex biochemical and integrated organ systems in the Trilobite, and for any increase in its information content, reliance on Darwinian evolution remains a matter of secular faith. We propose another theory of origins that involves an Intelligent Designer and Special Creation.
From:
Trilobites are extinct members of the large animal group (phylum Arthropoda) to which modern insects belong. They are well-represented in a long and detailed fossil record beginning in earliest Cambrian, 550 million radiometric years1 ago, and ending in the Permian, 250 million radiometric years ago. Universally they are found in the boundary between rocks relatively barren of metazoan life, and rocks containing abundant evidence of such life.
Trilobites are complex, elaborately
segmented forms with jointed appendages and swimmerets, antennae,  compound eyes,
and cephalized, or head-to-tail, nervous systems. Because they are extinct, very
little is known of their life habits except for deductions by association with
other forms that do have living representatives, and from careful examination of
the geologic deposits in which they are found. However, evolutionary theory
provides a conceptual framework for reconstructing the physiology and molecular
biology of this earliest widely distributed metazoan (complex, multicellular
organism).
compound eyes,
and cephalized, or head-to-tail, nervous systems. Because they are extinct, very
little is known of their life habits except for deductions by association with
other forms that do have living representatives, and from careful examination of
the geologic deposits in which they are found. However, evolutionary theory
provides a conceptual framework for reconstructing the physiology and molecular
biology of this earliest widely distributed metazoan (complex, multicellular
organism).
The tools of contemporary molecular systematics along with advances in understanding of molecular and cellular processes challenge the standard theory of undirected, naturalistic selection in Darwin's original proposal. Molecular features of a variety of organisms can now be compared, and genetic relationships, called phylogenetic linkages, can be constructed based upon those comparisons. With such powerful tools, it is not necessary to guess about operative processes in organisms no longer available for study. Thus, much of the molecular architecture of ancient organisms can be reconstructed with data readily available from contemporary living animals. The conclusions of such work are rather surprising.
Before continuing to explore the nature of the trilobite, a statement of premises employed in this reconstruction include:
Revealing the Past
The mechanisms operating in the trilobite's cells, tissues, and developmental processes when it first appeared on earth can be determined in precise detail.2
The assumption that complex molecular biological traits shared by disparate organisms require a shared ancestry is the basis of modern evolutionary taxonomy. Thus, molecular features shared by trilobites and mammals would require, at some time in the distant past, a common ancestor possessing those common features.3
Any other conclusion would assume highly unlikely events to have been repeated with exacting precision, falsifying the fundamental assumption of molecular systematics and taxing credulity beyond limits. Consequently, a complex feature shared by modern arthropods and humans, or arthropods and plants, was present in a common ancestor. A representation of such a proposed common ancestry for arthropods and humans is shown in Fig. 2.
Since trilobites were derived
arthropods, they too must have exhibited features shared by modern complex
animals, 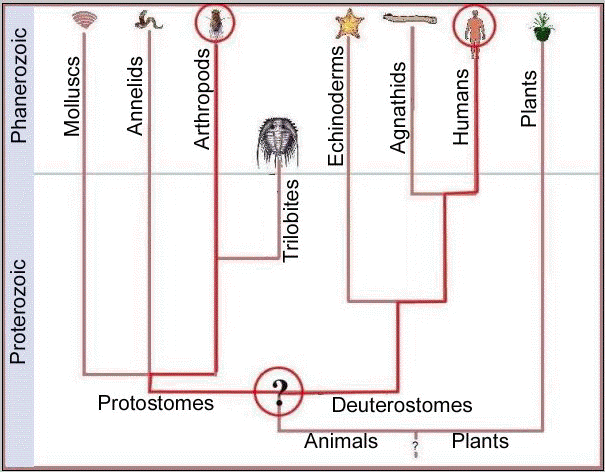 and we attribute complex features to this early metazoan with
confidence. Several examples are drawn from a large number of equally good
examples of complex molecular biological systems. It is necessary to include
some technical material in order to understand the level of complexity present
in cells, and the significance of this proposal.
and we attribute complex features to this early metazoan with
confidence. Several examples are drawn from a large number of equally good
examples of complex molecular biological systems. It is necessary to include
some technical material in order to understand the level of complexity present
in cells, and the significance of this proposal.
DNA and the Chromosome
Eukaryotic (nucleated) cells are the building blocks of all multicellular complex organisms, including humans. These cells are intricately constructed and highly integrated in function. The vast array of information in the cell is coded on long (up to 15 cm or more) molecular strands of DNA carried by the cell's chromosomes. Every somatic cell in the human body has a complement of 46 chromosomes; the total DNA of a single human cell would extend nearly two meters if stretched end-to-end. However, all of this material is housed in the cell nucleus, which has a diameter of about ten micrometers. This is a reduction by a factor of 200,000 in linear extension, a feat comparable to packing fifty miles of kite string into a shoe box.
The DNA must be organized very precisely to fit into the nucleus so that the cell has easy access to the genes, can carry out replication of entire strands, and can accurately divide replicas to daughter cells during cell division. This process is achieved by associating the DNA with a class of proteins called histones. Four different histones form a very stable octet containing two copies of each histone molecule. Since histones are positively charged to enable them to interact with negatively charged DNA, assembly of the octamer requires the aid of several special "scaffolding" proteins. This assembled histone core structure is so fundamental to cells that it is preserved across the entire spectrum of living eukaryotic cells almost without modification. For example, only one amino acid change distinguishes the histone H3 of a human from that of a sea urchin. Human H4 differs from H4 of a bean plant by only two amino acids, out of the hundred or so making up these proteins.
One and one-half turns of the DNA molecule (about 146 base pairs) are wrapped around each histone core to form a condensed structure called a nucleosome. These in turn are associated into higher order structures called solenoids, each a helical form comprised of six nucleosomes and a fifth histone protein, greatly reducing the overall extension of the chain. The solenoids are further condensed in a complex packing arrangement anchored to the backbone chromosome structure. The backbone is composed mainly of a class of proteins with remarkable properties that are attached to the DNA chain at specific sites. The protein can cut one strand of the double helix DNA structure at the point of attachment, hang on to the cut ends, pass the uncut strand through the cut ends, then rejoin the two ends, an operation that removes undesirable stresses from or adds desirable stresses to the chain. All these condensations reduce a 10 cm strand of DNA to an intricate structure 50,000-fold smaller in linear dimension. Because of its universal occurrence in all living things almost without variation, we can reasonably infer that this complex apparatus was also present in trilobite cells.
Cell Division
Without cell division there could be no growth in multicellular plants and animals. Before a cell can divide in a fashion that maintains its integrity and function, it must replicate its contents. The central core of biological memory coded in DNA must be duplicated so that an equivalent copy exists in each strand, producing another two meters of DNA in the case of human chromosomes. The two copies must then be separated from one another in such a way that one copy comes to reside in each daughter cell. To prevent dilution of cell contents, the cell must also make copies of all other molecules present and distribute these. This occurs in all eukaryotic cells, with essentially the same mechanisms. We will consider a few highlights of this incredibly complex process.
A human cell has 46 chromosomes to be duplicated. The ninety-two separate molecular assemblies must then move through the cytoplasm on directed journeys to the proper daughter cells. This process is mediated by microtubules, structural elements of the cell made up of a spiral array of protein molecules around a hollow interior space, much like a drinking straw. Chromosomes contain a special patch of protein where microtubules may attach, almost like a Velcro patch on fabric. When enough microtubules from opposite ends of the cell have attached to the two duplicated members of each chromosome pair, the chromosomes line up in the center, split apart, and the microtubules begin pulling their attached chromosomes through the cytoplasm to opposite ends of the dividing cell. The movement mechanism seems to involve contraction, expansion, and depolymerization of tubule structures as they pull, much in the fashion of tiny machines.4
This complex, elaborately controlled process is common to all eukaryotic cells and, therefore, presumably was already present in all essential details in the trilobite--one of the earliest metazoic fossils.
The Neuron and Synapse
The neuron, or nerve cell, transmits a nerve impulse to other neurons across the gap that lies between them, called a synapse, or synaptic gap. The resting neuron has a negative electrical potential on the inside of its membrane of about sixty millivolts. This potential is established by a special sodium/ potassium pump that uses cellular energy to pump positively charged sodium ions out of the cell. A nerve impulse is initiated and propagated by the movement of sodium ions back into the cell through special protein sodium channels in the membrane. Propagation is mediated by the successive opening of these channels, called voltage-gated sodium channels, along the length of the neuron's axon.
These channel proteins, located in neuron membranes, are intricately constructed. Each protein extends across the membrane twenty-four times, forming a barrel-shaped channel that has a voltage-sensitive gate. As the channel protein senses the depolarization of the nerve, the gate opens and sodium ions flow into the cytoplasm, propagating the voltage change and triggering the same response in adjacent channels. Once the membrane is fully depolarized, that is, the nerve impulse has passed, a "ball and chain" extension of the protein in the cytoplasm closes the channel, preventing further depolarization until the resting membrane potential has been reestablished by the sodium ion pump.
When an impulse reaches the terminus of a neuron, it must transfer the signal across the synaptic gap. In many cells, the transmission is effected by the release of a neurotransmitter substance, often acetylcholine, a small biomolecule. The acetylcholine accumulates in special membrane-bound synaptic vesicles within the cell. As a vesicle fills with neurotransmitter, it is transported through the cytoplasm toward the synapse by a unique protein called synapsin. This protein "walks" along microtubule highways of the cytoskeleton toward the membrane of the synaptic surface, carrying the synaptic vesicle along with it.
The vesicle membrane contains several proteins not found elsewhere in the cell. Two of these carry the technical names of synaptobrevin and synaptotagmin. Synaptobrevin binds a complex of proteins, which in turn binds to syntaxin, a protein in the cell membrane. Syntaxin thus anchors the acetylcholine-containing vesicle to the synaptic membrane. Synaptotagmin has two sites that can bind calcium ions. In the absence of calcium, synaptotagmin binds to the protein complex, preventing the vesicle from releasing its contents.
When an impulse reaches the synaptic region, calcium channels (similar to the voltage-gated sodium channels previously mentioned) are opened, allowing calcium to enter the cytoplasm. Synaptotagmin binds the calcium, allowing fusion protein to bind to the complex. In this state, the vesicle membrane can now fuse with the cell membrane, releasing acetylcholine into the synapse. The nerve impulse is thus transmitted, or propagated, to the neighboring neuron. All of these reactions of the traveling nerve impulse occur in milliseconds.
Because
this process represents
a very complex mechanism shared by insects and humans,
the reasonable assumption is that nerves and synapses in trilobites worked this
way also.
Cells contain many types of cytoplasmic vesicles. Each vesicle has, in addition to its normal component of membrane proteins, a special protein called Rab that directs a vesicle to its correct destination, much like a shipping label. These shipping labels are added when the vesicle is formed. They are "read" at the destination. If the vesicle has reached its proper site, it is retained. If not, it is redirected elsewhere. The synaptic vesicle must also have the correct label attached to be effective.
Other cytoplasmic proteins, called clathrin, identify an empty vesicle and surround it with a protein cage that preserves the membrane and the associated proteins from being lost. The empty vesicle remains enclosed in the clathrin cage until it has traveled away from the synaptic membrane into the cytoplasm for refilling.
This process, described in the barest details, is common to all animals with nervous systems from the simplest invertebrates to humans. Because this process represents a very complex mechanism shared by insects and humans, the reasonable assumption is that nerves and synapses in trilobites worked this way also. Thus, one of the earliest complex, mulicellular animals possessed the nervous system elements that are found in modern insects and humans.
Developmental Biology of Insects (and Trilobites)
Thanks to recent advances in understanding the molecular biology of development, a great deal can be inferred about the complex processes by which a single ovum in a mother trilobite becomes a functioning offspring. Trilobites belong to the same phylum as modern insects, so the corresponding formation of a typical metamorphosing insect, the fruit fly Drosophila, may be considered. Since these insects are very small, it is impractical for them to hatch a fully functional, winged offspring from a single, fertilized egg. The strategy of many insects is to lay an egg, which "hatches" into a stage called a caterpillar or larva. A larva is just a larger, developing "egg" with legs and a mouth for accumulating food material and eventually producing the adult form. Deep within the recesses of each caterpillar are the embryonic seeds of an entire adult organism. These special tissues, called imaginal disks, remain dormant until pupation, at which time the body of the caterpillar dissolves and the imaginal disks develop into the various parts of the adult. This is itself also a very complex process, but the sequence of events leading up to the formation of the imaginal disks gives remarkable insights into the complexity that can reasonably be concluded to be already present in trilobites.
While an insect egg is still in the ovary, unique distributions of special proteins are already being established within its cytoplasm. These proteins originate either from the egg nucleus or from maternal accessory cells surrounding the egg in the ovary. After fertilization, additional series of genes are activated, producing still other regulatory proteins in specific regions of the fertilized egg. The spatially asymmetric distribution of developmental proteins forms an early embryo in which each cell has a unique combination of regulators. The balance of these developmental gene regulators determines which genes are activated and which are suppressed in each cell. This asymmetry in turn determines head-to-tail, and other differentiation along the resulting body axis.
Genetic studies in Drosophila revealed that when developmental genes were mutated, they produced not just a single change such as eye color, but either massive and lethal effects or large changes in body form. For example, a single mutation in one such gene makes legs grow where antenna normally occur; another causes the formation of an extra body segment complete with extra wings. Extensive regulatory networks link activity for each of these developmental genes to hundreds of other more specific genes. Investigators have found that genes controlling fruit fly development are very similar in structure and action to those regulating vertebrate development, and that the genes often control analogous parts of the embryos of flies and humans. Thus, these developmental gene sequences, present in flies and humans, must also have been present in trilobites.
The
sequence of events
leading up to the formation
of the imaginal disks
gives remarkable insights
into the complexity
that can reasonably be concluded to be already present in trilobites.
Later studies have revealed the location of some of these genes on the Drosophila chromosome. When the main series of regulator genes (HOM-C genes) determining head-to-tail polarity and axial development in the Drosophila embryo was identified and mapped, investigators unexpectedly found that these lie on the chromosome in the same head-to-tail order as the portions of the anatomy whose development they control (colinearity). There is no obvious functional reason for this correlation of spatial arrangement to occur, and it would seem improbable if the organism's distinct parts had developed randomly at widely different times.
Even more recent studies have shown the existence of homologous types of regulatory genes responsible for ordering head-to-tail organization of the bodies of vertebrates, including humans. These genes, called Hox genes, are very similar to the corresponding genes in Drosophila (for some homeotic genes, the similarity between human and Drosophila is 98%), and they lie on the human chromosome in the same order as those found in the fruit fly. The inference of a common origin is very probable. Again, it is likely that this complex organization, along with the complexities already described for eukaryotic cell behavior, nerve synapse function, and all the other myriad, complex developmental and functional processes, were already in place in metazoan trilobites of the Cambrian, some of the earliest known multicellular forms.
The Trilobite Eye
The eye has been an object of wonder throughout recorded history because of its critical functions and complexity of organization. Recently discovered properties of some early trilobite eyes, which are similar to those found in modern insects, represent an "all-time feat of function optimization."5 The lens in each individual ommatidium, or facet, of the compound eye was composed of a single crystal of calcite (calcium carbonate) with the optical c-axis of the crystal coincident with the optical axis of the lens. This design presented an unusual problem for the trilobite, since a simple thick spherical lens of calcite could not have resolved light into a coherent image. These Lower to Middle Paleozoic trilobites, however, had a unique optical system unknown in any other creature that solves this problem.6
The optical system is a biconvex lens, composed of two lenses with differing refractive indices joined together. The interface of these two lenses is called a Huygens surface,7 as shown in Fig. 3.
The biconvex lens required an exact
shape for the trilobite eye to correctly focus light on receptors.8 In
Fig. 3, the left side shows how the incident light is focused into a coherent
image by the biconvex lens; the right side shows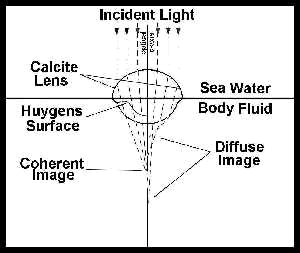 how the image is diffused
without the lens. Levi-Setti states:
how the image is diffused
without the lens. Levi-Setti states:
The realization that trilobites developed and used such devices half a billion years ago makes the shock even greater. And a final discovery--that the refracting interface between the two lens elements in a trilobite's eye was designed in accordance with optical constructions worked out by Descartes and Huygens in the mid-seventeenth century--borders on sheer science fiction.9
The significance of the biconvex lens of later trilobite ommatidia merits further elaboration, since a better example of intelligent design is hard to find. Levi-Setti continued: "When we humans construct optical elements, we sometimes cement together two lenses that have different refractive indices, as a means of correcting particular lens defects."10 Obviously, no one doubts that such optical elements in cameras, field glasses, and telescopes are intelligently designed, and Levi-Setti concurred in the case of the trilobite eye: "The design of the trilobite's eye could well qualify for a patent disclosure.11 When such a biconvex lens is found in nature, logic demands that intelligent design is a required element in the explanation of its formation.
Levi-Setti continued: "What we would like to hear, to appease our Darwinian upbringing,12 is that new visual structures were evolved in response to new environmental pressures as a means of survival."13 As a possibility he suggests that it "allowed the trilobite to see at some depth in sea, at dusk, or in turbid water.14" He added other imagined advantages, that they provided a prompter recognition and response to impending danger and that "mating may have proven more effective with sharper images."15
The earliest trilobites lacked the sophisticated lens described above, but had eyes that were apparently more like those of modern insects. No intermediate forms are known from the fossil record. When the Huygens lens is first found in trilobites, it was fully functional.
The regulatory mechanism of the early trilobite eye development must indeed be complex, since an estimated 2,500-5,000 genes appear to be involved in the developmental process of the insect eye.16 The ommatidium, or individual facet, of a compound eye such as that in Drosophila consists of a cluster of eight cells, seven of which develop into light receptors. One of these retinal cells, called R7, is responsible for detecting ultraviolet light (UV). Intensively studied for some years, the pathway from an undifferentiated cell to a sophisticated UV detector cell follows an intricate cascade of interactions.
The R7 cell membrane contains special proteins called receptor tyrosine kinase, or RTK. RTK projects active portions on either side of the membrane, both inside and outside the cell. The external RTK can join with another molecule of RTK to form a dimer in the presence of a specific activating agent. The internal portion of each RTK molecule then enzymatically attaches three phosphate groups to its partner. Thus phosphorylated, RTK binds a cellular protein, GRB2, and becomes activated to bind another protein, Sos. The Sos-GRB2-RTK complex can then interact with a critical membrane-bound protein called Ras. Ras is freed from the membrane into the cytoplasm, where it activates an enzyme called Raf. Activated Raf is able to bind another enzyme, MEK. MEK in turn activates a terminal cytoplasmic enzyme, MAP kinase, which apparently activates DNA-binding proteins and other key cellular proteins to change the direction of cellular differentiation. The cell can now become a normal R7 cell. Fundamentally similar processes are found in the cells of all multicellular eukaryotic organisms, presumably including the trilobite, and also (with slight differences) in the single-celled eukaryotes, yeast and protozoa.
The same
system
of genes
controlling eye development functioned
in the first trilobites.
Recently, manipulations in flies of a master eye developmental gene called Eyeless, caused the growth of eyes to be induced on wings, legs, and tips of the antennae. A similar master gene has been found in vertebrates, which have eyes that are completely different from insect eyes yet the developmental gene is nearly identical with that in Drosophila. When the appropriate gene from a mouse chromosome (presumably, the human gene would work as well) is inserted into a fly, it produces the specialized fly eyes wherever it is activated on the fly's body. The two genes are similar enough that the mammal gene can cause the formation of an insect eye. That line of reasoning then leads to the conclusion that the same system of genes controlling eye development functioned in the first trilobites.
More and more developmental pathways are found to be shared across a broad spectrum of organisms. Most of these would likely have been present in the trilobite. For example, genes responsible for the organization of the human front-to-back axis were discovered using the genes from Drosophila as molecular probes. Genes responsible for human brain organization in embryogenesis were discovered, again using Drosophila genes.
The eye, the hindbrain, the spinal cord, the pathing of axons, the differentiation of skeletal and heart muscle, the photoperiodic response, the sculpting of tissues involving select cell death (apoptosis), embryonic patterning, cell signaling, and a host of other "evolutionarily conserved" processes could also be cited as examples. The developmental gene called Hedgehog, directs the formation of limbs in fruit flies, while the equivalent vertebrate gene "Sonic Hedgehog" directs the formation of limbs in all known vertebrates, including human, mouse, chick, and fish. The elaborate control mechanisms precede any known organism with limbs, and all of these processes were probably operative in the trilobite.
The Problem of Complexity for One of the First Metazoa
Careful consideration has been directed to a few brief examples illustrating the complexity of living eukaryotic cells, nervous systems, developmental processes and organs already present in one of the first metazoa, the trilobite. These were drawn from among hundreds of other examples that could equally well have been used to make certain points. The trilobite, one of the earliest complex animals in the fossil record, first appeared in the Lower Cambrian.17 Trilobites are arthropods, in the same alliance as modern insects. The cells of trilobites divided in a manner similar to every modern eukaryote. The molecular mechanisms were all in place, all functioning as they do today in insects. The trilobites had nervous systems as complex as those of modern insects. The synapses in the nervous systems of trilobites functioned just as the synapses of all modern organisms do. The complex system of development of cephalized forms was already present and functioning. The eyes of trilobites manifest all of the complexity and developmental integrity of modern forms. Their eyes were developed by processes not only similar to those of other arthropods, but like those of vertebrates, including humans. A similar case could be presented for swimmerets and gills, legs, antennae, and intricately sculpted forms. Trilobites and all other forms appear on the scene as fully formed, fully competent organisms.
Since the complexities that have just been described were all present and fully functional in one of the first multicellular animals for which there is a record, the questions may be asked: Where did these complexities come from? Where and when did evolution take place? There is no indisputable evidence of any earlier form from which they could have been derived. Conway Morris, in discussing steps that might have led up to the true arthropods (the phylum to which trilobites belong), concluded: "All this is very speculative indeed, and really raises more questions than it answers."19 Furthermore, there is no evidence for the existence of a mechanism in biological systems for adding information to complex systems. To argue that they came from Precambrian forms which were not preserved because they had no hard parts, is to argue from the absence of evidence. Fossils with preserved soft parts have in fact been found in the Cambrian and Upper Precambrian sediments in many localities.20 There is no confirmed Precambrian evolutionary sequence leading up to the trilobite that can be accounted for by Darwinian mechanisms, hence the conclusion that there was no Precambrian Darwinian evolution of trilobites.
Conclusion
The complex biochemical systems and the integrated organ systems of the trilobite just described did not happen by accident. Darwinian mechanisms have not been demonstrated to be active, causal factors, nor have they been universally considered a reasonable scientific explanation of the phenomena described in this paper. When it has been tried, the result is no more effectual than the attempt proposed by Levi-Setti to account for the formation of the trilobite's double lens. Studies of these systems of the trilobite are an indictment of the inadequacies of Darwinian evolutionary theory. This may be why some evolutionary authors, when they write books about the earliest life forms, carefully skirt the sudden appearance of endlessly complex forms. Their attitude seems to be: "If it's there, evolution must be able to do it."
Yet we have seen from a careful consideration of the evidence that the origin of the complex biochemical systems and integrated organ systems of trilobites, and by extension, biological organisms in general, cannot be accounted for by Darwinian evolution (an extension of a naturalistic philosophy, in which there is no role for a Creative Intelligence). When Darwinian evolution is used as an explanation for the existence of complex living systems, it becomes a philosophical or even a quasi-religious view held by those who wish the world to have no Designer.
While design and its purposes can be reasonably inferred from biological data presented in this paper and while design logically implies the operation of an intelligent designer, the designing agent and mechanism employed by the designer cannot be identified by scientific means. We have few if any conceptual tools at this time with which to pin down the elusive concept of intelligence in biology so that it can be studied and characterized. The mode, mechanism, locus, domain, or modus operandi of intelligence in biology is not known. The interface between intelligence and the material world remains a mystery at this time.
At least two possible approaches to studying intelligent design can be explored. First, one may posit that there is a purposive or functional logic embedded in living organisms that is as real and objective as the laws of physics. The nature and origin of this logic and its role in effecting change in the biological world would be the focus of study. This approach places design squarely in the natural order.
The second approach may be to grant the above, but to claim further that the designer acted in nature throughout the history of life in the universe to bring about certain purposes. This approach would acknowledge that recognizing an Intelligent Designer and the mode of operation lies outside the competence of science and must be approached through interdisciplinary methods and concepts of theology and philosophy. Indeed, when one pushes beyond the design inference and its purposes, one leaves the domain of science and enters that of philosophy/theology. The purposes of the Intelligent Designer would be the focus of study, and their effect on the history of life in the universe would be considered. Clearly, this is an area ripe for further study by scientists, philosophers, and theologians within the Judeo-Christian tradition.
While the authors also believe that intelligent design originates in the mind of a Supreme Intelligent Designer, the Christian God, and is actualized as part of God's purpose for the universe, this belief is not essential to the study of Intelligent Design as a scientific theory. Design is a reasonable inference, as witness the trilobite, and as such is amenable to scientific study by believer and unbeliever alike.
©2000
Acknowledgment
The authors wish to thank Joy Chadwick for her indispensable assistance in preparing the manuscript for publication.
Notes
1Radiometric means the measurement of geologic time based on the disintegration of radioactive elements.
2The rationale for arguing that studies of molecular features of present day animals can tell us about the molecular features of ancient organisms is supported by Harold J. Morowitz, a biophysicist, who wrote about the very earliest protocells. He says: "In practice, the outputs of metabolism have a universal as well as a specific character ... Within the universal core of the metabolic chart, we can assume we are recovering knowledge of 3.7 billion-year-old or older biochemistry ... we are studying the biochemistry of the universal ancestor" (Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis [New Haven: Yale University Press, 1992], 51).
3J. S. Levinton, G. Wray, and L. Shapiro, "Molecular Evidence for a Deep Precambrian Divergence of Animal Phyla. I. Introduction and Regression Approach," Geological Society of America annual meeting, Denver, CO, Abstracts with Programs, 28:7 (1996): A-52.; G. Wray, J. S. Levinton, and L. Shapiro, "Molecular Evidence for a Deep Precambrian Divergence of Animal Phyla. II. Relative Rate Tests and Implications," Ibid.; and G. A. Wray, J. S. Levinton, and L. H. Shapiro, "Molecular Evidence for Deep Pre-Cambrian Divergences Among Metazoan Phyla," Science 274 (1996): 568-73.
4David J. Sharp, Gregory C. Rogers, and Jonathan M. Scholey, "Microtubule Motors in Mitosis," Nature 407 (September 7, 2000): 41-47.
5The nuclear physicist Riccardo Levi-Setti (Director of the Fermilab at the University of Chicago) and trilobite authority has made an in-depth study of trilobite eyes, reported in his book, Trilobites, 2d ed. (Chicago: The University of Chicago Press, 1993).
6Levi-Setti, Trilobites, 29-74.
7The optical principles utilized by this trilobite were elaborated by Huygens (and Descartes) in the seventeenth century, but the trilobite lens worked perfectly well using these optical principles long before the Dutch mathematician figured out how.
8Levi-Setti, Trilobites, 55.
9Ibid., 54.
10Ibid., 44.
11"Ibid., 57.
12Levi-Setti's honesty is admirable. By his own admission, it is his personal development that directed him to seek an evolutionary explanation. Rather than seeking a new explanation he tried to "shoehorn" the data into an evolutionary framework.
13Ibid., 59.
14Ibid.
15Ibid., 66.
16G. Rubin, "Secrets in the Fly Eye," Discover 17:7 (July 1996): 110.
17 The basal Cambrian is sometimes loosely defined as the point in the geologic column where the first trilobites appear.
18S. Conway Morris, The Crucible of Creation (New York: Oxford University Press, 1998), 184.
19L. Spetner, Not By Chance (New York: Judacia Press, 1997)
20S. Bengston and Y. Zhao, "Fossilized Metazoan Embryos from the Earliest Cambrian," Science 277 (September 12, 1997): 1645-48; S. Xiao, Y. Zhang, and A. H. Knoll, "Three-dimensional preservation of algae and animal embryos in Neoproterozoic phosphorite," Nature 391 (February 5, 1998): 553-57; C.-W. Li and J.-Y. Chen, "Cambrian Sponges with Cellular Structures," Science 279 (February 6, 1998): 879-82.
Additional References
E. Boncinelli, A. Simeone, A. La Volpe, H. Faiella, D. Acampora, and L. Scotto, "Human cDNA Clones Containing Homeobox Sequences," Cold Spring Harbor Symposium, Quantitative Biology 50 (1985): 301-6.
J. P. Grotzinger, S. A. Bowring, B. Z. Saylor, and A. J. Kaufman, "Biostratigraphic and Geochronologic Constraints on Early Animal Evolution," Science 270 (1995): 598-604.