 Science
in Christian Perspective
Science
in Christian Perspective
Science
in Christian Perspective
Is "Progressive Creation"
Still a Helpful Concept?
John Jefferson Davis*
Gordon-Conwell Theological Seminary
130 Essex Street, S. Hamilton, MA 01982
From Perspectives on Science and Christian Faith 50 (December 1998): 250-259.
This article argues that Bernard Ramm's concept of "progressive creation" is still a useful category for interpreting biblical and scientific data relating to origins. After reviewing significant developments since Ramm's 1954 publication in both theological and scientific scholarship, an attempt is made to relate a modified version of Ramm's concept to current discussions of creation and evolution in English-speaking Christianity. It is argued that "progressive creation," which can be distinguished from both "fiat creation" and "theistic evolution," is a category broad enough to encompass both the immanent presence of God working within nature and the transcendent power of God above the laws of nature. Christians working in the life sciences are left free to recognize the variety of ways in which the creative action of God has been operative in the natural order.
In the estimation of one recent historian of science, Bernard Ramm's 1954 book, The Christian View of Science and Scripture, "...profoundly influenced the way in which many orthodox Christians answered the questions posed by creation and evolution."1 Twenty-five years after the book's publication, John W. Haas, Jr., a member of the American Scientific Affiliation, an association of evangelicals in the natural and biological sciences, called Ramm's book "a pivotal event" in the modern history of science and religion.2
In his discussion of the biological sciences, Ramm proposed the concept of "progressive creation" as an alternative to both theistic evolution and the "fiat creationism" of the fundamentalists, who understood creation almost exclusively in terms of instantaneous, supernatural acts of God. According to Ramm, progressive creation was the means by which the Spirit of God, as "World Entelechy," brought to pass the divine will in nature. Under the direction of the Holy Spirit, the laws of nature, over a long period of time and through a variety of processes, actualized the plan of God.3 Because he believed in several acts of sudden, fiat creation in the history of the earth-in view of the discontinuities in the fossil record-Ramm believed that his view was clearly differentiated from theistic evolution. In Ramm's mind, "progressive creation" avoided the arbitrariness of fiat creationism, while preserving its emphasis on the transcendence of God, and also avoided the "uniformitarianism" of theistic evolution, while affirming its emphasis on progress and development.4
Ramm expressed the rather grandiose and somewhat naive hope that his concept of "progressive creation" could form the basis of "a new biological synthesis" which would be for biology what the relativity theory was for physics.5 Quite contrary to Ramm's intentions, his proposal sparked a renewal of the "flood geology" and "young-earth creationist" tradition in American fundamentalism - a tradition which Ramm had hoped to lay to rest.6
The purpose of this essay is to argue that "progressive creation" is still a useful category for interpreting the biblical and scientific data relating to origins. Of course, much has transpired both in biblical scholarship and scientific research since Ramm published a generation ago. After reviewing significant developments in the theological and scientific areas, I will relate a modified version of Ramm's concept of "progressive creation" to current discussions of creation and evolution in English-speaking Christianity.
Trends in Biblical Scholarship
In recent years, biblical scholars have increasingly drawn attention to the fact that the agenda which modern interpreters often brought to the text of the early chapters of Genesis - issues of "science and Scripture"- are, at best, secondary to the primary interests of the biblical writers. Evangelical scholars, such as Gordon Wenham, stress that Genesis is to be seen primarily against the background of its ancient Near Eastern religious environment. According to Wenham, Genesis 1-11 is best seen as a "tract for the times," challenging the ideas of the polytheistic religions of the ancient Near East about the nature of God, the world, and humankind. Genesis is concerned with affirming the unity of God in the face of polytheism and the justice of God rather than caprice; "scientific" issues in the modern sense of the word are only indirectly related to the primary purpose of the text.7
Similarly, Gerhard Hasel has argued for the "polemic nature of the Genesis cosmology."8 A primary concern of the text is to criticize the polytheistic nature religions of the ancient Near East which identify the sun, moon, stars, and forces of nature as deities. The Genesis cosmology forcefully asserts that the heavenly bodies are not gods and goddesses to be worshiped, but creatures of the one, holy God who created the world and rules it according to his righteous laws. The Genesis cosmology represents a complete break with the mythological cosmologies of Israel's neighbors in the Fertile Crescent.9
Shortly after Ramm published The Christian View, the evangelical theologian J. I. Packer made the valuable observation that interpreters must draw distinctions between "...the subjects about which the Scripture speaks and the terms in which it speaks of them." The writers of Scripture spoke about the natural world in an ordinary, nontechnical language shared with their contemporaries. Their concern was not primarily the inner structure of the world and of humans, but with the relationship of both to God.10
This point concerning the "relational" rather than the "scientific" focus of Genesis has also been made in another way by British geneticist R. J. Berry. Since the end of the eighteenth century, in Berry's view, liberal and conservative interpreters alike have erred in approaching the biblical accounts of creation "as if they were primarily concerned with origins rather than with relationships."11 The preoccupation with questions of origins has deflected attention away from the primary biblical concerns, namely, the relationship of nature to God and humanity's proper relationship to the creation. As a result, evangelicals have failed to develop an adequate theology and practice of environmental stewardship.
Discussions of "creation and evolution" have at times suffered from a lack of attention to the range of possible meanings of the biblical terms used to describe God's creative work. For example, the primary sense of the crucial word "bara" ("create"), used 49 times in the Old Testament and ten times in Genesis, is that through God's command something comes into being that had not existed before. The word is used exclusively of divine action, and implies a creative work that is beyond human power. Since the word never occurs with the object of the material, the primary emphasis of the term is on the newness of the created object. The concept of "ex nihilo" creation, while it may be implied in a given case, is not necessarily inherent in the meaning of the word.12
It is important to note that the focus of such a biblical term for creation as bara is not so much on the physical processes used by God as on the results of the divine action and the relationship of these results to the redemptive purposes of God. Modern science is primarily concerned with physical processes; Scripture is primarily concerned with results and relationships13 The word bara can be used to refer to a clearly supernatural, ex nihilo creative act which brings the universe into being (Gen. 1:1), or to the divine power working through natural processes to "create" the winds (Amos 4:13), or to bring animals to birth through the normal processes of gestation (Ps. 104:30). Whether God's creative work in any given case involves natural or supernatural means, or long periods of time rather than instantaneous effects, cannot be judged in advance, but must be determined in light of the particular biblical texts and specific features of the natural order. This distinction between process and results will be noted again in the further reflections on "progressive creation" and "theistic evolution" in the closing section of this paper.
Scientific Developments since Ramm
The biological sciences have developed dramatically since Ramm wrote The Christian View of Science and Scripture in 1954.14 Some of the most explosive growth has been in the disciplines of genetics and molecular biology. The discovery of the double- helix structure of the DNA molecule by Watson and Crick in 1953 and the subsequent deciphering of the genetic code were watershed events in the history of biology, opening up new frontiers of scientific research.15 Biologists were able to study living forms not only externally, but internally as well, at the genetic and molecular levels.
According to Stephen Jay Gould, the most important event in evolutionary biology during the 1970s was the "development of electrophoretic techniques for the routine measurement of genetic variation in natural populations."16 These new techniques allow biologists to compare the sequences of the bases in the chains of genetic material (DNA and RNA) and the amino acid sequences of proteins which are characteristic of each organism. The quantitative differences in these sequences are interpreted as a measure of the degree of the remoteness of two organisms from a common ancestor. These discoveries in genetics and molecular biology provide new ways for biologists to check hypotheses about the relationships between living forms previously limited to studies of morphology and embryology.17
During the last twenty years, paleontologists have made important new discoveries, especially in the fossil record of the vertebrates. New groups of jawless fish, sharks, amphibians, and dinosaurs have become known, and major transitions between amphibians and reptiles, reptiles and mammals, and dinosaurs and birds have been extensively studied.18
In the 1970s two paleontologists, Niles Eldredge of the American Museum of Natural History and Stephen Jay Gould of Harvard University, sparked a major controversy in evolutionary biology and paleontology with their concept of "punctuated equilibria." According to this view, evolution proceeds in "fits and starts" rather than in small, gradualistic Darwinian steps. Evolution proceeds very rapidly when new species are being formed, and then these forms typically remain unchanged for long periods of time ("stasis"). According to the "punctuationists," the well-known gaps in the fossil record are real, and are to be expected if speciation occurs rapidly in small, geographically isolated populations that would leave few fossil remains.19 Defenders of the orthodox Darwinian gradualism argue that the "punctuationists" have overstated their case. The coarse time resolution of most fossil studies biases the observer toward a perception of stasis. Some cases seem to fit the "punctuationist" model fairly well, but other groups, e.g., the mammals, seem to have adapted and changed in very gradual and piecemeal ways.20
As late as 1953, when paleontologist George Gaylord Simpson published his classic book on macroevolution entitled The Major Features of Evolution,21the major evolutionary transitions between the larger taxonomic groups were still, for the most part, inadequately documented from the fossil record.22 Darwin himself was quite aware of the major "gaps" in the fossil strata and attributed them to the "Imperfections" of the geological record.23 Since the 1950s, new fossil discoveries have given evidence of some transitional forms that are clearly intermediate between fish and amphibians and between reptiles and mammals.
Beginning in 1952, Jarvik and other paleontologists have given
extensive attention to a primitive extinct amphibian, Ichthyostega, discovered in
the sedimentary rocks of eastern Greenland. Ichthyostega had anatomical features
which were intermediate between the more advanced amphibians and the crossopterygian
(lobe-finned) lung fish from which amphibians are believed to have evolved. The extinct
lung fish, Eusthenopteron and Ichthyostega (see Fig. 1 below), share many
common features, including the ability to breathe air, the shape of the body, and the
skeletal features of the upper limbs and skull. Ichthyostega had a genuine
fish-like tail, its skin was protected by small fish-like scales, and its compressed body
shape was typical of fish. Yet in spite of its strange mixture of fish and amphibian
characteristics, Ichthyostega was clearly a full-fledged tetrapod and can be placed
among the early amphibians, of which it is a primitive representative.24
Fig. 1
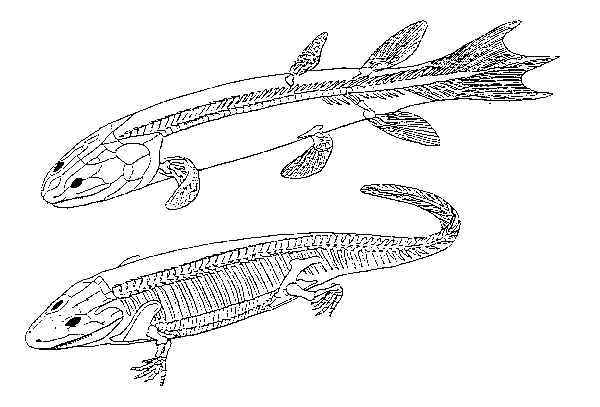
A comparison of the skeletons of the lobe-finned fish Eusthenopteron
(above) and the primitative amphibian Ichthyostega (below).
Reprinted with permission from Colbert and Morales, Evolution of the Vertebrates,
4th ed., (New York: John Wiley a& Sons, 1991), 69.
Even more recently Per Ahlberg, a paleontologist working at the Natural History Museum in London, has reported new findings related to Panderichthys, an extinct lobe-finned fish. His findings show that Panderichthys had a mosaic of fish-like and amphibian-like characteristics.25 These fossils indicate that changes in the skull roof, braincase, and fins occurred in a relatively rapid period (geologically speaking) of 9-14 million years, and provide further evidence of the transition between the strictly aquatic lobe-finned fish and the four-legged amphibians.26
The fossil sequence from the reptiles to the earliest mammals "...is the most fully documented of the major transitions in vertebrate evolution," according to Robert Carroll, a paleontologist at McGill University.27 The transformations in the fossil record can be traced over a period of 150 million years, from "small, cold-blooded scaly reptiles to tiny, warm-blooded, furry mammals."28 Here it is a case not of "gaps in the fossil record," but rather an abundance of riches. T. S. Kemp of the University Museum and Department of Zoology at Oxford concurs: "This is the one known example where the evolution of one class of vertebrates from another class is well documented by the fossil record."29
Most paleontologists agree that the ancestry of the mammals is to be sought among a group of extinct mammal-like reptiles known as cynodonts.30 The fossil record of mammal-like reptiles (synapsids) is the most complete of any group of terrestrial vertebrates with the exception of the mammals themselves.31 By way of illustration, we may examine the case of Cynognathus, an extinct cynodont about the size of a large dog, displaying a blend of reptilian and mammalian characteristics (see Fig. 2 below).
Fig 2
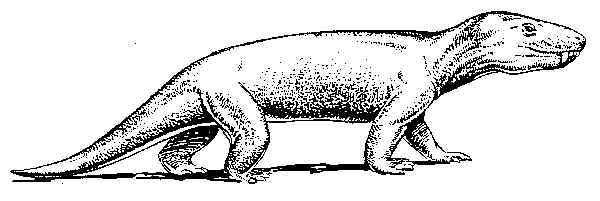
Cynoganthus, an extinct mammal-type reptile.
Reprinted with permission from Colbert and Morales, Evolution of the Vertebrates,
4th ed., (New York: John Wiley a& Sons, 1991), 126.
Cynognathus had a rather large skull that was dog-like in appearance. Its teeth were differentiated and specialized, unlike the undifferentiated teeth of a reptile. Small, peg-like incisors were adapted for biting and nipping, and the post-canines were suited for chewing food - an indication that this animal cut its prey into small pieces before eating it, rather than swallowing it whole as do many reptiles. The vertebral column was differentiated into cervical, dorsal, and lumbar regions. To increase the efficiency of locomotion, the limbs were held beneath the body with the knee pointing forward and the elbow pointing backward. These and other specializations of Cynognathus show that it was an active, carnivorous reptile that was approaching a mammalian stage of development in many respects.32 Cynognathus is only one example from a very large class of extinct tetrapods that display characteristics that are intermediate between the reptiles and the mammals.
Other living forms-the monotremes, or egg-laying mammals of Australia-also exhibit features intermediate between reptiles and the more advanced mammals. The echidna and the platypus or duckbill (see Fig. 3) are very primitive mammals that reproduce by laying eggs and suckle their young on milk secreted by modified sweat glands.
Fig 3


Platypus (above) and Echidna (below).
Reprinted with permission from Colbert and Morales, Evolution of the Vertebrates,
4th ed., (New York: John Wiley a& Sons, 1991), 240.
The skeletons and soft anatomies of these animals display certain reptilian characteristics33 The cervical ribs are not fused, and certain reptilian characteristics can be seen in the skull. The urogenital system and rectum open into a common cloaca as in reptiles rather than separately as in mammals. In many respects, monotremes, such as the platypus and the echidna, give, according to Colbert and Morales, excellent living examples of "mammals intermediate in their stage of evolution between the mammal-like reptiles and the higher mammals."34
Any discussion of intermediate forms in the geological record must include the fossil evidence of the hominid forms believed to be the precursors of modern Homo sapiens.35 Much new evidence has become known since Ramm wrote in 1954. The hominid fossil record shows that the anatomical and behavioral characteristics that we associate with the "human" have emerged and developed over long periods of time. For example, the Australopithecines, extinct hominids that flourished in Africa approximately 3-4 million years before the present, were capable of upright walking, like modern humans, but in brain capacity and other anatomical features were more like chimpanzees. The Australopithecines in these respects are intermediate forms between modern man and the great apes. As the hominid fossil record is followed over a four-million-year period from the Australopithecines to Homo habilis to Homo erectus to Homo neanderthalensis to modern Homo sapiens, one can see the changes in brain capacity, dentition (tooth structure), and skeletal structures that move from forms that are more apelike to those more characteristic of modern humans. This evidence of transitional forms in the fossil record, together with evidence from comparative anatomy and molecular biology,36 leads modern biologists and anthropologists to believe that modern humans and the great apes shared a common ancestor some five million years ago, probably on the African continent.
One of the most notable features of the fossil record is the sudden, almost "explosive" appearance of the major phyla during the late Precambrian and early Cambrian periods. Since then extensive changes have occurred within the phyla, but few new animal phyla have appeared. It is also quite notable that, according to Eicher and McAlester: "There is no fossil record of the origin of these phyla, for they were already clearly separate and distinct when they first appeared as fossils."37 Complex, multicellular organisms such as the trilobites, corals, and crustaceans appear fully formed in the fossil record with no obvious ancestral forms.
Various explanations have been offered for the "Cambrian explosion," which has been called the "Big Bang" in the history of life on earth. Some scientists have alleged that increases in oxygen levels in the oceans could have promoted the rapid development of life. Other suggestions have included increases in the calcium or phosphorous content of the oceans, enabling the rapid development of organisms with skeletons; the advent of predators providing selection mechanisms for diversification; and the evolution of regulatory genes making the rapid appearance of new body plans possible.38 All these suggestions are rather speculative, and no single explanation has won general acceptance within the scientific community.
Before 1947 almost nothing was known about the nature of multicellular life prior to the Cambrian. During that year an extensive fossil deposit of soft-bodied organisms, dating to about 640 Myr before present, was discovered in the Ediacara Hills of southern Australia. These fossils, which have since become known as the "Ediacara fauna," fall into four main categories: jellyfish, soft corals, segmented worms, and other organisms of unknown affinities (see Fig. 4).39 These animals did not possess shells or skeletons.
Fig. 4
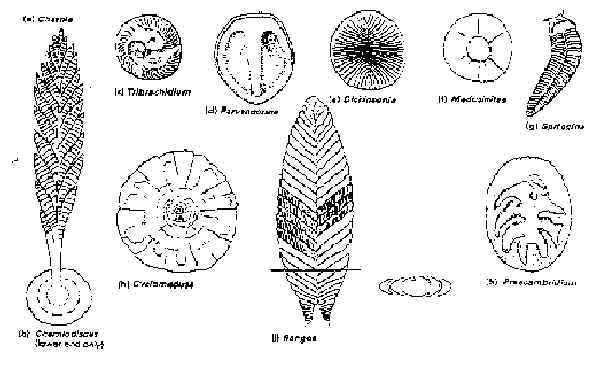
Precambrian Edicara fauna.
From Clarkson, Invertebrate Palaeontology and Evolution, 2nd. ed. (London:
Allen & Unwin, 1986), 49.
Reprinted with kind permission from Kluwer Academic Publishers
In recent years, a rich and varied fauna from the lower Cambrian-Precambrian boundary has been discovered in Siberia. These tiny fossils, many of which have phosphatic shells, are generally known as the "Tommotian fauna" (see Fig. 5). Similar fossils have since been found in Australia, England, and Scandinavia. Many of these forms continue up into the lower Cambrian, where they appear with the trilobites.40
Fig 5
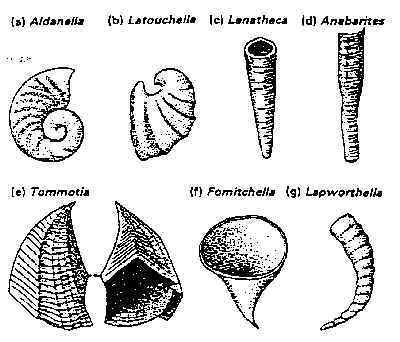
Tommotian fauna from Siberia.
From Clarkson, Invertebrate Palaeontology and Evolution, 2nd. ed. (London:
Allen & Unwin, 1986), 49.
Reprinted with kind permission from Kluwer Academic Publishers
It is not clear how either the soft-bodied Ediacara fauna or the small, shell-bodied Tommotian fauna could plausibly be seen as ancestral to complex organisms such as the trilobites. In the words of E. N. K. Clarkson of the University of Edinburgh, these fossils testify to "an explosive development of life at this most critical point in life's history."41
Some of the most extensively studied animals from the Cambrian period are the trilobites, hard-shelled creatures somewhat like modern horseshoe crabs, that are abundantly represented in the fossil record from the lower Cambrian, 570 Myr before present, until their final extinction during the Permian, about 250 Myr ago. The trilobites appear in the fossil record abruptly and fully formed. Their origins are a major mystery in the history of life. According to H. B. Whittington, a leading authority in this field, "unequivocal answers cannot be given" to the questions of how and where the trilobites arose; no transitional series of fossils have been found.42 Candidates for the presumed ancestors of the trilobites have been found among the annelid worms or the Ediacara genus Spriggina (see Fig. 4), but as Clarkson has admitted, "this is only speculation."43
Since the 1960s, scientists have extensively studied the remarkable compound eyes of the trilobites (see Fig. 6).44 They are the most ancient visual system known in the entire history of life. Like the eyes of insects and crustaceans, these are compound eyes composed of radially arranged visual units that give a wide-angled visual field. The number of optical elements in these compound eyes could range from about one hundred to more than fifteen thousand in a single eye. It is believed that with such eyes the trilobite could form an image of a nearby object and even estimate its distance. The problem, from a purely evolutionary perspective, is how to explain the origin of such a complex and abruptly appearing organ through a process of small, gradual changes and natural selection. No living forms prior to the trilobites give evidence of having even rudimentary eyes, much less complex eyes like those of the trilobites.
Fig. 6
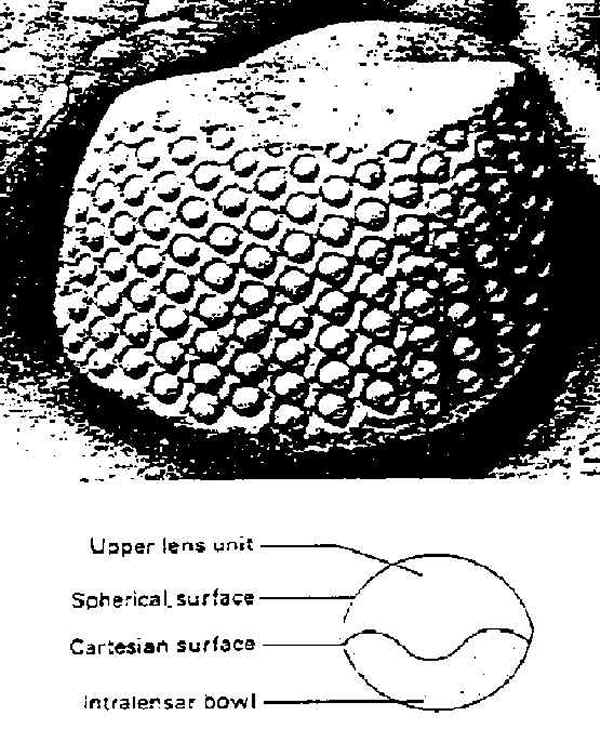
Trilobite eyes and their structure.
Reprinted with permission from R. Boardman, ed., Fossil Invertebrates,
Oxford: Blackwell Science Ltd., 1987), 227
When Ramm published The Christian View, experimental research on the origins of life was in its infancy. In 1953 Stanley Miller and his associates at the University of Chicago had just completed the first successful prebiotic simulation experiments, synthesizing a variety of amino acids by passing electric sparks through a mixture of methane, ammonia, hydrogen, and water vapor.45 Since Miller's pioneering work, four decades of intensive chemical experimentation have shown that the synthesis of amino acids is rather easy to achieve, but the prebiotic synthesis of the more complex molecules necessary for life is extremely difficult.46 The basic problem facing origins-of-life researchers is that life requires the presence of very complex molecules (proteins, enzymes) for metabolism, and very complex molecules (RNA, DNA) for replication, and these molecules must occur together47 The enormous difficulties of creating such molecules under conditions similar to those obtaining in the early earth led Francis Crick, the co-discoverer of the structure of the DNA molecule, to propose the bizarre hypothesis ("directed panspermia") that life was sent to earth by an advanced civilization in the form of bacteria on a spaceship.48
Some leading researchers in this field believe that RNA was the original molecule in the origins of life, somehow initially serving both as a metabolic catalyst and as an informational "template" for replication. Chemist Manfred Eigen has synthesized an RNA molecule with the aid of an enzyme extracted from some living bacteria, and Leslie Orgel has synthesized RNA from simpler molecules using a form of RNA as a template. But neither Eigen nor Orgel have been able to synthesize RNA without the presence of either an enzyme or a template, as would have been the case under the actual conditions of the prebiotic earth.49
Other researchers, such as A. G. Cairns-Smith, a chemist at Glasgow University, believe that the problems of life originating in the primeval ocean are overwhelming, and have proposed as an alternative the "clay hypothesis." According to Cairns-Smith's speculations, the first organisms on earth had a different biochemistry from that of life today, and evolved through natural selection from inorganic crystals. Complex molecules were built up on a substrate of clay, which in some fashion was presumably able to supply the information needed for replication now provided by RNA and DNA.50 In one recent experiment, researchers were able to build up long molecules on mineral surfaces, but these molecules were not able to replicate themselves, as in a living system.51
It would seem that the creation of life in the laboratory, if it is indeed possible, is a long way in the future. While investigators have proposed many models for the origins of life, Leslie Orgel, a leading authority in the field, has admitted that "...evidence in favor of each of them is fragmentary at best." In a somewhat somber prognostication, Orgel concludes: "The full details of how ... life emerged may not be revealed in the near future."52
Some Concluding Reflections
The foregoing survey has shown that momentous discoveries have occurred in the life sciences since Bernard Ramm wrote over a generation ago. Nevertheless, it remains my conviction that Ramm's concept of "progressive creation" is still a helpful way of interpreting both the biblical and scientific data relating to origins. "Progressive creation," understood as an alternative to "fiat creation" and Atheistic evolution" incorporates the elements of truth in both, and means that God's creative action has occurred over long periods of time through a variety of means.
The emphasis on "a variety of means" calls attention to the fact that the focus of the biblical terminology of creation is on the results of God's action, and the relationship of those results to the divine purpose, rather than on the details of the processes used by God to achieve these results. "Fiat creationism" in both its older and more recent forms in American fundamentalism is based on an unnecessary dichotomy between natural and supernatural processes as possible methods of creation.53 God is free to create through natural or supernatural means, and by rapid processes or over long periods of time; no single type of process can, in an a priori fashion, be identified as uniquely suited to the divine purpose.54
"Progressive creation," like "theistic evolution," recognizes that a Christian theory of origins must acknowledge and incorporate the evidence for the evolutionary changes that have occurred in the history of life. In some cases, such as the reptilian-mammalian transitions in the fossil record noted above, the evidence for macro-evolutionary change is stronger than Ramm supposed, and "theistic evolution" would seem to be an appropriate terminology.55 In other cases, however, the evidence for discontinuity and the rapid emergence of novelty in the history of nature is strong, and the language of "progressive creation" calls attention to these facts56 Notable cases in point here would be the ex nihilo creation of space-time, matter, and energy at the "Big Bang"; the emergence of life from inanimate matter over 3.5 billion years before the present; the explosive appearance of the major animal phyla at the Precambrian-Cambrian boundary some 570 million years ago; and the sudden appearance of art and other expressions of behaviorally modern humanity, some forty thousand years ago.
The terminology, "progressive creation," is broad enough to encompass both the immanent presence of God working within the laws of nature and the transcendent power of God above the laws of nature. From this perspective, the Christian working in the life sciences is free to recognize the variety of ways in which God's creative activity has been expressed, in the confidence that the "book of nature" rightly interpreted will ultimately be consistent with the "book of Scripture" rightly understood.
�1998
Notes
1Ronald L. Numbers, The Creationists (Berkeley: University of California Press, 1992), 187.
2John W. Haas, Jr., "The Christian View of Science and Scripture: A Retrospective Look," Journal of the American Scientific Affiliation 31 (1979): 117.
3Bernard Ramm, The Christian View of Science and Scripture (Grand Rapids: Eerdmans, 1954), 116.
4Ibid., 113, 116.
5Ibid., 272.
6Numbers, The Creationists, 187f. Numbers traces the responses by John C. Whitcomb, Jr., Henry Morris, and the launching of the modern "creation science" movement, signalized by the 1961 publication of Whitcomb and Morris, The Genesis Flood, defending (against Ramm) a young earth, a literal six-day creation, and a universal flood as the causal explanation for the earth's major geological features.
7Gordon Wenham, Genesis 1-15 (Waco, TX: Word, 1987), xlv, 1.
8Gerhard F. Hasel, "The Polemic Nature of the Genesis Cosmology," Evangelical Quarterly 46 (1974): 81-102. The ancient Near Eastern religious context of Genesis, and ancient religious ideas of creation, are masterfully reviewed in Claus Westermann, Genesis 1-11: A Commentary, trans. John J. Scullion (London: SPCK, 1984), 19-46, "Creation in the History of Religions and in the Bible." Further helpful discussion of the background and theological purposes of the creation accounts in Genesis may be found in Bruce K. Waltke, "The Creation Account in Genesis 1:1-3: Part I: Introduction to Biblical Cosmogony," Bibliotheca Sacra 132 (1975): 25-36.
9Hasel, "The Polemic Nature of the Genesis Cosmology," 91.
10J. I. Packer, Fundamentalism and the Word of God (London: InterVarsity Fellowship, 1958), 96-8.
11R. J. Berry, "Creation and the Environment," Science and Christian Belief 7 (1995): 21-43 at 21. The references in this article provide a comprehensive review of historical and theological literature relating to environmental issues.
12H. Ringgren, "bara," Theological Dictionary of the Old Testament, Vol. II, eds. G. Johannes Botterweck and Helmer Ringgren, trans. John T. Willis (Grand Rapids: Eerdmans, 1975), 242-9; and Thomas E. McComiskey, "bara," Theological Wordbook of the Old Testament, Vol. I, ed. R. Laird Harris (Chicago: Moody Press, 1980), 127-8.
13In terms of Aristotle's terminology of the four types of causes, modern science, in its concern with the quantitative understanding of physical processes, focuses on "material" and "efficient" causes; the Bible and Christian theology, with its concern for the meaning and purpose of God's creative work, focuses on "formal" (issues of design, designer) and "final" (issues of purpose, teleology) causes.
14Surveys of current trends in evolutionary biology may be found in standard texts such as Douglas J. Futuyma, Evolutionary Biology, 2d ed. (Sunderland, MA: Sinauer Associates, 1986) and Mark Ridley, Evolution (Boston: Blackwell Scientific Publications, 1993). Historical studies of the development of modern biological thought are presented in Ernst Mayr, The Growth of Biological Thought: Diversity, Evolution, and Inheritance (Cambridge, MA: Harvard University Press, 1982) and David Young, The Discovery of Evolution (Cambridge: Cambridge University Press, 1992). Young traces the historical development of evolutionary thought from the seventeenth century to the present against the background of the growth of the disciplines of botany, zoology, and geology, in a clearly written and helpfully illustrated text.
15For the historical development of the discipline of genetics and reviews of basic concepts, see John B. Jenkins, Genetics (Boston: Houghton Mifflin, 1975); George W. Burns and Paul J. Bottino, The Science of Genetics, 6th ed. (New York: Macmillan, 1989); David Suzuki and Peter Knudston, Genetics: The Clash Between the New Genetics and Human Values (Cambridge, MA: Harvard University Press, 1989); and Gunther Stent, Molecular Genetics: An Introductory Narrative (San Francisco: W. H. Freeman, 1971). D. J. Weatherall, The New Genetics and Clinical Practice, 3d ed. (Oxford: Oxford University Press, 1991) is considered the definitive work on genetics and clinical ethics. The impact of new genetic research on evolutionary biology is discussed in Christopher Wills, The Wisdom of the Genes: New Pathways in Evolution (New York: Basic Books, 1989). For the original discovery of the structure of the DNA molecule, see James D. Watson, The Double Helix: A Personal Account of the Discovery of the Structure of DNA, ed. Gunther S. Stent (London: Weidenfeld and Nicolson, 1981).
16Stephen Jay Gould, Ontogeny and Phylogeny (Cambridge, MA: Harvard University Press, 1977), 406.
17Colin W. Stearn and Robert L. Carroll, Paleontology: The Record of Life (New York: John Wiley and Sons, 1989), 29.
18Robert L. Carroll, Vertebrate Paleontology and Evolution (New York: W. H. Freeman and Co., 1988), preface. For readable introductions to the field of paleontology, see Stearn and Carroll, op. cit.; Steven M. Stanley, Earth and Life Through Time (New York: W. H. Freeman, 1986) - combines historical geology and paleontology; and Richard Fortey, Fossils: The Key to the Past (London: Heinemann, 1982) - well- illustrated.
19The case for "punctuated equilibria" is argued by Steven M. Stanley, The New Evolutionary Timetable: Fossils, Genes, and the Origin of Species (New York: Basic Books, 1981), and by Niles Eldredge, Time Frames: the Rethinking of Darwinian Evolution and the Theory of Punctuated Equilibria (London: Heinemann, 1986). This latter work includes as an appendix (pp. 193-223), the original 1972 paper by Eldredge and Gould, "Punctuated Equilibria: an Alternative to Phyletic Gradualism."
20effrey Levington, Genetics, Paleontology, and Macroevolution (Cambridge: Cambridge University Press, 1988), 407-8. The orthodox, neodarwinian "gradualist" paradigm is also defended by Antoni Hoffman, Arguments on Evolution: a Paleontologist's Perspective (New York: Oxford University Press, 1989). It should be noted that this debate is an intramural dispute within evolutionary biology, not an abandonment of the evolutionary paradigm itself.
21G. G. Simpson, The Major Features of Evolution (New York: Columbia University Press, 1953).
22Robert L. Carroll, "Revealing the Patterns of Macroevolution," Nature 381 (2 May 1996): 19-20 at 19. Simpson and Carroll were aware, of course, of the famous series of horse fossils documented by Marsh and others in the nineteenth century, and the notable Archaeopteryx, intermediate between the dinosaurs and birds, but had in mind the relative absence of forms clearly intermediate between major groups such as the fish and amphibians, and the reptiles and the mammals.
23The awareness of major discontinuities in the fossil record was a significant factor in the development of the "punctuated equilibria" hypothesis by Eldredge and Gould. Today, many paleontologists would prefer to describe the fossil record as "biased" rather than as "inadequate." They would point out that the process of fossilization is inevitably biased in favor of the preservation of hard-bodied rather than soft-bodied organisms, and in favor of those animals that die near large, shallow bodies of water rather than at higher elevations, and so on. When these biases are taken into account, paleontologists "... can make corrections for them in order to arrive at a balanced view of the life of the past" (Stearn and Carroll, Paleontology, 12).
24This information on Ichthyostega and the fish-amphibian transition is from Edwin H. Colbert and Michael Morales, Evolution of the Vertebrates: A History of the Backboned Animals Through Time, 4th ed. (New York: John Wiley & Sons, 1991), 67-9; I. I. Schmalhausen, The Origin of the Terrestrial Vertebrates, trans. Leon Kelso (New York: Academic Press, 1968), 34 ff.; and Robert L. Carroll, Vertebrate Paleontology and Evolution (New York: W. H. Freeman, 1988), 158 ff.
25Per E. Ahlberg, Jennifer A. Clack, and Ervins Luksevics, "Rapid Braincase Evolution between Panderichthys and the Earliest Tetrapods," Nature 381 (2 May 1996): 61-3.
26Carroll, "Revealing the Patterns of Macroevolution," 20.
27Carroll, Vertebrate Paleontology and Evolution, 361.
28Ibid., 362.
29T. S. Kemp, Mammal-Like Reptiles and the Origin of Mammals (London: Academic Press, 1982), 1. Kemp's statement implies, of course, that the transitions between other major vertebrate classes are less well-documented and based on inferences from small numbers of fossil remains.
30Colbert and Morales, Evolution of the Vertebrates, 228.
31Kemp, Mammal-Like Reptiles and the Origin of Mammals, 1.
32The above description is from Colbert and Morales, Evolution of the Vertebrates, 123-5.
33Ibid., 241.
34Ibid.
35The hominid fossil evidence is reviewed and analyzed in works such as Michael H. Day, Guide to Fossil Man, 4th ed. (Chicago: University of Chicago Press, 1986); Richard G. Klein, The Human Career: Human Biological and Cultural Origins (Chicago: University of Chicago Press, 1989); Roger Lewin, Human Evolution: An Illustrated Introduction, 3d ed. (Boston: Blackwell Scientific Publications, 1993); Ian Tattersall, The Fossil Trail (New York: Oxford University Press, 1995); and John Reader, Missing Links: The Hunt for Earliest Man (New York: Penguin Books, 1988).
36Anatomically, modern humans are more similar to the great apes (the gorillas and chimpanzees) than to any other living forms. At the molecular level of DNA sequences, humans and chimpanzees share an approximately 98% degree of similarity.
37Don L. Eicher and A. Lee McAlester, History of the Earth (Englewood Cliffs, NJ: Prentice-Hall, 1980), 236. The beginning of the Cambrian is generally dated approximately 570 million years (Myr) before the present.
38T. Peter Crimes, "The Period of Evolutionary Failure and the Dawn of Evolutionary Success: the Record of Biotic Changes across the Precambrian-Cambrian Boundary," in Stephen K. Donovan, ed., The Paleobiology of Trace Fossils (New York: John Wiley & Sons, 1994), 105-33 at 125. Richard Forty, Fossils: The Key to the Past (London: Heinemann, 1982) has noted: "The acquisition of shells and skeletons is one of the great milestones in the history of the biosphere, and the difficulty of finding a single neat explanation only adds to the fascination" (p. 148).
39E. N. K. Clarkson, Invertebrate Paleontology and Evolution, 2d ed. (London: Allen & Unwin, 1986), 48.<P7MJ247>
40Ibid., 51-2.
41Ibid., 52.
42H. B. Whittington, Trilobites (Woodbridge, UK: Boydell Press, 1992), 84-5.
43Clarkson, Invertebrate Paleontology and Evolution, 331.
44See Riccardo Levi-Setti, Trilobites: A Photographic Atlas (Chicago: University of Chicago Press, 1975), 23 ff.; Richard S. Boardman, ed., Fossil Invertebrates (London: Blackwell Scientific Publications, 1987), 227 ff.; and Whittington, Trilobites and Clarkson, Invertebrate Paleontology and Evolution above. According to Levi-Setti, the recently discovered properties of the trilobite eye lenses "represent an all-time feat of function optimization" (p. 23); the trilobites had apparently in some remarkable fashion "discovered" and applied sophisticated principles of optics scientifically described by Descartes and Huygens in the seventeenth century so as to be able to correct for spherical aberration of light and form sharp images (p. 38).
45These and subsequent experiments are described in Stanley L. Miller and Leslie Orgel, The Origins of Life on the Earth (Englewood Cliffs, NJ: Prentice Hall, 1974), 83-102. Influential in origins of life research were the earlier speculations of Russian scientist A. I. Oparin, The Origin of Life on the Earth, 3d ed., trans. Ann Synge (Edinburgh: Oliver and Boyd, 1957). Oparin, operating from the philosophical standpoint of Marxist-Leninist "dialectical materialism," attributed self-organizing properties to matter: "Matter never remains at rest, it is constantly moving and developing...[it] changes from one form of motion to another...each more complicated and harmonious than the last. Life thus appears as a particularly very complicated form of the motion of matter, arising as a new property at a definite stage in the development of matter" (p. xii). While not necessarily sharing Oparin's dialectical materialism, other workers in this area, recognizing the great improbability of the chance origins of life, have similarly attributed powers of "self-organization" to inanimate matter.
46A very helpful overview and analysis of origins of life research is provided by Freeman Dyson, Origins of Life (Cambridge: Cambridge University Press, 1985), esp. 1-34.
47Metabolic processes are those by which the living organism converts nutrients from its environment into useful forms of energy; "replication," governed by the RNA and DNA molecules, refers to the processes by which the cell duplicates itself and its substructures.
48Francis Crick, Life Itself: Its Origin and Nature (London: Macdonald & Co., 1981). Needless to say, Crick's suggestion has not been taken very seriously by the scientific community.
49On the work of Manfred Eigen, see Manfred Eigen, Steps Toward Life: A Perspective on Evolution, trans. Paul Woolley (Oxford: Oxford University Press, 1992), and Manfred Eigen, et al., "The Origin of Genetic Information," Scientific American 244:4 (April 1981): 78-94; for a good overview of Orgel's work and a review of research in the field, see Leslie E. Orgel, "The Origin of Life on the Earth," Scientific American 271:4 (October 1994): 53-61.
50A.G. Cairns-Smith, Genetic Takeover and the Mineral Origins of Life (Cambridge: Cambridge University Press, 1982); a simplified layperson's account of this theory is presented in A. G. Cairns-Smith, Seven Clues to the Origin of Life (Cambridge: Cambridge University Press, 1985). Cairns-Smith believes that among the intractable problems faced by the more popular "primeval soup" model are: contamination of early prebiotic molecules by other chemical substances; the difficulty of achieving sufficiently strong concentrations of the essential molecules in the primordial ocean; and the problem of hydrolysis: left to themselves, complex molecules in a watery solution would break down into the simpler constituent amino acids (Genetic Takeover, 45-59).
51James P. Ferris, Aubrey R. Hill, Jr., Rihe Liu, and Leslie Orgel, "Synthesis of Long Prebiotic Oligomers on Mineral Surfaces," Nature 381 (2 May 1996): 59-61.
52Orgel, "The Origin of Life on the Earth," 61.
53For example, Henry Morris and Gary Parker, representing the "creation science" point of view (young earth, six-day creation, "Flood geology") state: "Evolution purports to explain the origin of things by natural processes, creation by preternatural process; and it is semantic confusion to try to equate the two" (Morris and Parker, What Is Creation Science? [El Cajon, CA: Master Books, 1987], 300). This would seem to be an example of the logical fallacy known as the fallacy of the excluded middle: "X must be explained in terms [and only in terms] of either A or B." Rather, it may be the case that X can be explained by C or D, or by some combination of A, B, C, D, and so forth. In the case of origins, it needs to be recognized that God is free to create through either natural or supernatural means, or through a combination of both.
54Rather than a two-fold distinction between "natural" and "supernatural" means, it is more biblically accurate to recognize a three-fold distinction among God's works of ordinary providence, special providence, and miracle. In ordinary providence, God works immanently through the regular laws of nature (e.g., causing the grass to grow through the processes of photosynthesis [Ps. 104:14]; creating animals through the normal processes of gestation [Ps. 104:24, 30]); in extraordinary providence, God redirects the forces and laws of nature (e.g., causing a wind to blow quail from the sea to feed the Israelites during the wilderness wanderings [Num. 11:31]); in miracles God transcends the laws of nature for a redemptive purpose (e.g., the floating axhead, [I Kings 6:6]; the feeding of the 5000; and the bodily resurrection of Jesus).
55In the perspective being argued here, "theistic evolution" is understood as a subcategory within the larger framework of "progressive creation." Biblical texts such as Gen. 1:20, 21, where God is said to create mediately by addressing the waters and the earth, can be seen as consistent with creation through natural processes. Biblical texts such as Gen. 1:21, 24 ("kinds") and I Cor. 15:39 ("all flesh is not the same": birds, animals, fish) can be seen as statements concerning the results of God's creative work - the major groups of animals are distinct and not interfertile - rather than excluding descent from a common ancestor with modification as a possible process of origination.
56Some proponents of theistic evolution as the more inclusive category seem to be reticent to recognize special divine interventions in the natural order. Howard J. Van Till, for example, believes that the creation's "functional integrity" and the natural order's "God-given creaturely capacities" are "...sufficiently robust so as not to require additional acts of special creation...to actualize the full array"...of life forms that have ever existed" (Van Till, "Basil, Augustine, and the Doctrine of Creation's Functional Integrity," Science and Christian Belief 8:1 [1996]: 21-38 at 29). See also Van Till, The Fourth Day: What the Bible and the Heavens are Telling Us about the Creation (Grand Rapids: Eerdmans, 1986), esp. chaps. 1-5. The terminology of "progressive creation" argued in the present article seems to be a more adequate way of explicitly affirming the theological categories of miracle (e.g., the resurrection) and special providence (e.g., answers to petitionary prayer) that are essential for biblical faith.