
On The Origin of Stasis By Means of Natural Processes
Arthur L. Battson III
7668 Dartmoor Avenue
Goleta, CA 93117
From: PSCF 46 (December 1994): 230-241. Response: Krause Bossard
p. 230
"I am well aware that there is scarcely a single point discussed in this volume on which facts cannot be adduced, often apparently leading to conclusions directly opposite to those at which I have arrived. A fair result could be obtained only by fully stating and balancing the facts on both sides of each question, and this cannot possibly be done here." (Charles Darwin, The Origin of Species, 1859)
Charles Darwin was well aware that scientists could come to directly opposite conclusions from those set forth in his Origin of Species. Although his theory could account for minor evolutionary change and the diversity of finches, Darwin knew that he had to virtually ignore the natural history of life on earth in order to maintain any hope of accounting for the origin of the phyla and the major disparity between arthropods and anthropologists.
Darwinian theory is in conflict with the most prominent features of earth's natural history. First of all, geology does not provide the transitional forms Darwin's theory demands. In 1859, the conflict with paleontology was the most serious objection to the theory and over the years the gap between data and theory has only grown wider. Today scientists acknowledge fewer transitional forms than Darwin thought existed. Species typically arise suddenly and "fully formed."
The second conflict between Darwinism and natural history is the phenomenon of stasis. Geology reveals the stability of forms rather than their gradual transformation into substantially different body plans. The stability of the higher taxa in particular suggests the existence of natural processes which prevent major evolutionary change from occurring on a gradual step-by-step basis.
Finally, the pervasive pattern of geologic succession is systematically backwards from that predicted by the theory. Darwinian theory predicts that the gradual accumulation of minor evolutionary change and the increasing diversity of the lower taxa should ultimately produce the profound differences among the major body plans and the disparity of the higher taxa. Diversity should precede disparity. Geologic succession reveals the opposite: disparity precedes diversity. The major themes or body plans appear suddenly in the history of life only to be followed by variations on these pre-existing themes. The natural history of life on earth is systematically top to bottom, not bottom to top as Darwinian theory predicts.
Had Darwin developed a theory to explain the empirical data of natural history, he should have come to directly opposite conclusions. He should have developed a theory to explain why species do not gradually transform into substantially different body plans on a gradual step-by-step basis. The phenomenon of stasis and the stability of the major body plans is based upon an abundance of data and our theories describing the natural world should explain that data. The empirical evidence suggests the need to develop a theory which is based upon natural history rather than one which must explain away its key features. Although neo-Darwinian theory helps to explain minor evolutionary change, a theory of "macrostasis" needs to be developed that explains the stability of the major body plans.
p. 231
Conflicts Between Darwin and Paleontology
Darwin saw evolution as a slow and stately process. He pictured organisms gradually transforming from one species into another over immense spans of time. Evolution, he believed, had to occur through "infinitely numerous transitional links" forming "the finest graduated steps." Darwin was a strict adherent of gradualism and the notion that "nature does not make leaps." He spelled this out very clearly in his Origin of Species:
If it could be demonstrated that any complex organ existed which could not possibly have been formed by numerous successive slight modifications, my theory would absolutely break down (Darwin, 1859, p. 219).
There was one major stumbling block to this view of life: the fossil evidence. In a chapter entitled "On the Imperfection of the Geological Record," he readily admits:
... The number of intermediate varieties, which have formerly existed on the earth, (must) be truly enormous. Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps, is the most obvious and gravest objection which can be urged against my theory (Darwin, 1859, p. 292).
Despite the serious problems the geologic evidence presented, Darwin believed that the passage of time would reveal the enormous number of transitional forms his theory demanded. However, such was not to be the case. David Raup, former curator of geology at Chicago's Field Museum of Natural History, put it this way:
Well, we are now about 120 years after Darwin, and knowledge of the fossil record has been greatly expanded ... ironically, we have even fewer examples of evolutionary transition than we had in Darwin's time. By this I mean that some of the classic cases of Darwinian change in the fossil record, such as the evolution of the horse in North America, have had to be discarded or modified as a result of more detailed information ... (Raup, 1979).
The paleontological case against gradualism was serious in Darwin's day and time has only made matters worse. Stephen Jay Gould, professor of geology and paleontology at Harvard University, explains:
The history of most fossil species include two features particularly inconsistent with gradualism:
1) Stasisñmost species exhibit no directional change during their tenure on earth. They appear in the fossil record looking much the same as when they disappear; morphological change is usually limited and directionless;
2) Sudden appearanceñin any local area, a species does not arise gradually by the steady transformation of its ancestors; it appears all at once and `fully formed' (Gould, 1977).
Gould honestly admits that the neo-Darwinian synthesis is not supported by the fossil evidence and "is effectively dead, despite its persistence as textbook orthodoxy" (Gould, 1980).
Although stasis is the dominant feature of the history of life, exceptions to the general pattern of stasis can be cited. Examples of transitional series can be found at lower taxonomic levels. At higher taxonomic levels, however, transitional sequences range from scarce to non-existent. Evidence of gradualism between phyla, classes, and even orders is either non-existent or is much disputed. Certainly, no pervasive pattern of gradualism exists.
p. 232
George Gaylord Simpson acknowledged this decades ago as he described the situation in these terms:
This is true of all thirty-two orders of mammals...The earliest and most primitive known members of every order already have the basic ordinal characters, and in no case is an approximately continuous sequence from one order to another known. In most cases the break is so sharp and the gap so large that the origin of the order is speculative and much disputed...
This regular absence of transitional forms is not confined to mammals, but is an almost universal phenomenon, as has long been noted by paleontologists. It is true of almost all classes of animals, both vertebrate and invertebrate...it is true of the classes, and of the major animal phyla, and it is apparently also true of analogous categories of plants (Simpson, 1944).
Recent research on the origin of the higher taxa confirms what paleontologists have known for decades.
Taxa recognized as orders during the (Precambrian-Cambrian) transition chiefly appear without connection to an ancestral clade via a fossil intermediate. This situation is in fact true of most invertebrate orders during the remaining Phanerozoic as well. There are no chains of taxa leading gradually from an ancestral condition to the new ordinal body type. Orders thus appear as rather distinctive subdivisions of classes rather than as being segments in some sort of morphological continuum (Valentine, Awramik, Signor, and Sadler, 1991).
The origin of classes and phyla constitute an even greater difficulty for neo-Darwinian theory. Compounding the problem is the small window of time available for the origin of the vast majority of phyla. Recent research has squeezed "Biology's Big Bang" down to a few million years. (See Kerr, 1993 and Bowring et al., 1993.) When one compares the period of time it has taken Darwinian processes to modify the beak of a finch with the period of time virtually all of the major body plans appeared in the Cambrian explosion, it becomes difficult to believe that a gradual accumulation of microevolutionary changes had much at all to do with the origin of the higher taxa.
Darwinian evolution predicts the regular presence of transitional forms. The fossil record reveals their regular absence. It also reveals a natural phenomenon which, until recently, was virtually ignored by paleontologists. That phenomenon is stasis. The tragedy of Darwinism is that it has impeded the progress of science by turning the attention of biologists and paleontologists away from the empirical data and distracting them from developing theories which explain the pervasive natural phenomenon of stasis. For over 130 years, scientists working within the Darwinian paradigm have attempted to develop theories to explain data which, on the macro level, do not exist (See Figure 1).
 |
|
Fig. 1 Darwinian Theory vs. the Fossil Record Darwinian theory attempts to explain the
common ancestry of all |
Writing in the introduction to the 1956 reissue of the Origin of Species, W.R. Thompson commented:
The success of Darwinism was accompanied by a decline in scientific integrity. This is already evident in the reckless statements of Haeckel and in the shifty, devious and histrionic argumentation of T. H. Huxley...
To establish the continuity required by the theory, historical arguments are invoked even though historical evidence is lacking. Thus are engendered those fragile towers of hypotheses based on hypotheses, where fact and fiction intermingle in an inextricable confusion (Thompson, 1956).
The fossil data clearly show patterns of stasis rather than of major evolutionary sequences and it is this phenomenon to which scientists must turn their attention. As Niles Eldredge and Stephen Jay Gould put it: "Stasis is data" (Gould, 1991). Scientists cannot afford to lose sight of this abundant historical evidence.
p. 233
Gould recently described the importance of understanding stasis in these terms:
...we must understand that nothing happens most of the time ó and we don't because our stories don't admit this theme ó if we hope to grasp the dynamics of evolutionary change. (This sentence may sound contradictory, but it isn't. To know the reasons for infrequent change, one must understand the ordinary rules of stability.) The Burgess Shale teaches us that, for the history of basic anatomical designs, almost everything happened in the geological moment just before, and almost nothing in more than 500 million years since (Gould, 1988).
It is entirely conceivable that natural processes alone are insufficient to overcome what Gould has referred to as "the ordinary rules of stability." Kurt Wise, a former doctoral student of Gould, has suggested that there might be at least four distinct levels of stasis: molecular-level, population-level, species-level, and higher taxon-level stasis. Although Wise believes that the first three levels of stasis are violable, he points out that there may be a mechanism preventing change in higher taxa which is inviolable. Rejecting Gould's metaphysical assumptions, Wise concludes that natural processes probably exist which prevent major evolutionary change from transforming the baramin, or originally created kinds, into significantly different body plans:
It is probably only the stasis on the level of higher taxa which is both valid and differs qualitatively from the other levels of stasis. Only higher taxa lack demonstrable evidence of change...Higher taxon-level stasis could conceivably be the result of what might be called Baraminic Stasis ó the permanent constraint of organisms under natural conditions to stay within the bounds of their baramin (Wise, 1991).
The concept of the baramin is synonymous with the concept of the "created kind" and is anathema to scientists who believe that the origin and diversity of life must be attributed to purely mechanistic pro-cesses. The goal of science, however, should not be to develop a naturalistic "creation account" in an attempt to explain the origin and diversity of all life by purely materialistic means. Instead, the goal of science should be to most accurately describe the pervasive patterns and phenomena found in nature even if those natural processes prevent major evolutionary change from occurring. Science needs a diversity of ideas unencumbered by philosophical naturalism. Scientists need a theory to explain the phenomenon of higher taxon-level stasis and a theory to explain why species do not appear to gradually evolve into something substantially different.
Conflicts Between Darwin and Geological Succession
Prior to the time The Origin of Species was written, the geologic time scale in its modern form was already fully developed. In fact, the time scale based on fossils was built by geologists who were creationists (Raup, 1981). Scientists of Darwin's day did not equate geologic succession with evolution, nor should we today. One reason is that geological succession does not reveal how new species came into existence, it only reveals when. Another reason is that the order of appearance found in the fossil record is, as a general rule, systematically backwards from the major predictions of Darwinian theory.
Geological succession is often looked upon as primary evidence for the fact of evolution. Amoebas appear before fish, and fish before philosophers. In the popular view, this succession from simple to complex is evidence for evolution. Geological succession, however, does not necessarily proceed from simple to complex. For example, trilobites, among the most advanced of the arthropods, are the first arthropods to appear in the geological record. Darwin lamented over the complexity of the vertebrate eye, confessing that
to suppose that the eye...could have been formed by natural selection, seems, I freely confess, absurd in the highest possible degree (Darwin, 1859, p. 217).
Yet some scientists believe that the schizochroal eyes of some trilobites possessed the most sophisticated optical systems ever utilized by any organism (Levi-Setti, 1975). David Raup has stated that the trilobites
...used an optical design which would require a well trained and imaginative optical engineer to develop today... (Raup, 1979).
The early metazoans were anything but simple. The complexity of the early metazoans, however, is but a wrinkle in the irony of Darwinism. The real irony lies in the general systematic order of appearance of the taxa in the fossil record.
Life is far too complex for our theories to predict the specific body plans which should come into existence as the result of evolution. Both Darwinian gradualism and punctuated equilibrium, however, predict that a systematic branching pattern should be evident if all life has arisen from a common ancestor (see Figures 2 and 3). Both of these models are based upon relatively small intra- or inter-species change and, therefore, predict that numerous transitional species must come into existence which gradually diverge to produce the disparity of the higher taxa.
|
|
|
Fig. 2
Phyletic gradualism |
p. 234
Both models predict that as new species emerge and morphological distance between them increases, new genera and eventually new families should appear. As increasing diversity occurs, new orders, new classes, and ultimately new phyla should come into existence. In short, diversity should precede disparity. The gradually increasing diversity of the lower taxa should ultimately result in the disparity of the higher taxa and the appearance of major new body plans. The pattern of geological succession predicted by both of these
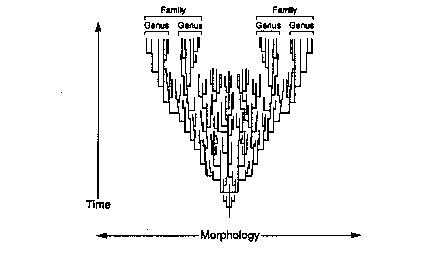 |
|
Fig. 3 Punctuated Equilibrium |
evolutionary theories is from bottom to top: species to genera to families to orders to classes to phyla.
The pervasive pattern of geological succession is contrary to these theories (see Figure 4). Disparity precedes diversity. The initial appearance of virtually all phyla occurs with very low species diversity. The origin of the major body plans is not the result of the increasing diversity of the lower taxa; the general pattern is not bottom to top. Rather, the dominant pattern is top to bottom, contrary to theory. As paleontologists Douglas Erwin, James Valentine, and John Sepkoski describe the situation:
The fossil record suggests that the major pulse of diversification of phyla occurs before that of classes, classes before that of orders, and orders before families. This is not to say that each higher taxon originated before species (each phylum, class, or order contained at least one species, genus, family, etc. upon appearance), but the higher taxa do not seem to have diverged through an accumulation of lower taxa (Erwin, Valentine, and Sepkoski, 1988).
In another article, Valentine and Erwin review hypotheses regarding the mode of origin of animal body plans to
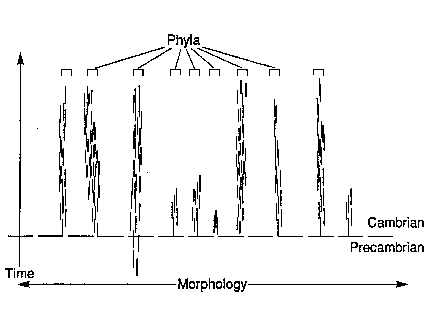 |
|
Fig 4. The Fossil Evidence |
determine their consistency with the fossil evidence. They conclude that both Darwinian gradualism and punctuated equilibrium are inadequate to account for the appearance of invertebrate body plans and their major modifications:
The models we consider are of three sorts: those that extrapolate processes of speciation to account for higher taxa via divergence, those that invoke selection among species, and those that emphasize that many higher taxa originated as novel lineages in their own right, not only as a consequence of species-level processes. It is in this latter class of model that we believe the record favors (Valentine and Erwin, 1985, p. 71).
If large populations have gradually evolved, there should be unmistakable evidence in the fossil record, yet it is simply not found.
...many of the large populations should have been preserved, yet we simply do not find them. Small populations are called for, then, but there are difficulties here also. The populations must remain small (and undetected) and evolve steadily and consistently toward the body plan that comprises the basis of a new phylum (or class). This is asking a lot. Deleterious mutations would tend to accumulate in small populations to form genetic loads that selection might not be able to handle. Stable intermediate adaptive modes cannot be invoked as a regular feature, since we are then again faced with the problem of just where their remains are. We might imagine vast arrays of such small populations fanning continually and incessantly into adaptive space. Vast arrays should have produced at least some fossil remains also. Perhaps an even greater difficulty is the requirement that these arrays of lineages change along a rather straight and true course ó- morphological side trips or detours of any frequency should lengthen the time of origin of higher taxa beyond what appears to be available. Why should an opportunistic, tinkering process set on such a course and hold it for so long successfully among so many lineages?
p. 235
We conclude that the extrapolation of microevolutionary rates to explain the origin of new body plans is possible, but does not accord with the primary evidence (Valentine and Erwin, 1985, pp. 95, 96).
The model of punctuated equilibrium or species selection attempts to account for the lack of evidence by relying primarily on the evolution of small isolated populations which would have a diminished chance of leaving a fossil record. This scenario has its difficulties, however, as Valentine and Erwin point out:
The required rapidity of the change implies either a few large steps or many and exceedingly rapid smaller ones. Large steps are tantamount to saltations and raise the problems of fitness barriers; small steps must be numerous and entail the problems discussed under microevolution. The periods of stasis raise the possibility that the lineage would enter the fossil record, and we reiterate that we can identify none of the postulated intermediate forms. Finally, the large numbers of species that must be generated so as to form a pool from which the successful lineage is selected are nowhere to be found. We conclude that the probability that species selection is a general solution to the origin of higher taxa is not great, and that neither of the contending theories of evolutionary change at the species level, phyletic gradualism or punctuated equilibrium, seem applicable to the origin of new body plans (Valentine and Erwin, 1985, p. 96).
This evidence further substantiates the proposition that minor lower-level evolutionary change cannot be extrapolated to account for major evolutionary change. This appears to be true for the both the tortoise and the hare, Darwinian gradualism and punctuated equilibrium. The tortoise is far too slow to account for the fossil evidence and the hare spends far too much time in stasis.
Darwin admitted that the geological evidence was
the "most obvious and gravest objection which can be urged against my
theory."
Darwin admitted that the geological evidence was the "most obvious and gravest objection which can be urged against my theory." At the time, he was primarily concerned with the lack of transitional forms in the fossil record. Today, those concerns are compounded by fewer transitional forms than Darwin had in his day, and by the systematically upside-down order of geological succession. Darwin was ironically prophetic in stating that the facts can lead to conclusions directly opposite to those at which he arrived.
Roger Lewin further describes the origin of most major body plans in the Science Research News report, "A Lopsided Look at Evolution":
Described recently as "the most important evolutionary event during the entire history of the Metazoa," the Cambrian explosion established virtually all the major animal body forms ó Bauplane or phyla ó that would exist thereafter, including many that were 'weeded out' and became extinct. Compared with the 30 or so extant phyla, some people estimate that the Cambrian explosion may have generated as many as 100. The evolutionary innovation of the Precambrian/Cambrian boundary had clearly been extremely broad: "unprecedented and unsurpassed," as James Valentine of the University of California, Santa Barbara, recently put it (Lewin, 1988).
p.236
Lewin then asked the all important question: "Why, in subsequent periods of great evolutionary activity when countless species, genera, and families arose, have there been no new animal body plans produced, no new phyla?" (Lewin, 1988). If neo-Darwinian theory is true, why should the Cambrian contain a greater number of body plans than exist today, particularly with such low species diversity? Figures 5 and 6 graphically illustrate the situation.
 |
|
Fig 5. The Cambrian Explosion |
Lewin's question leads us to an even more important question. What mechanisms have prevented major evolutionary change from occurring over the past 500 million years? Why did the origin of the phyla appear to have stopped first, followed by classes and then orders?
 |
|
Fig. 6 The Present |
One rather convincing explanation that has been offered for the pattern is based upon the fact that any major novelty that arises relatively quickly through non-selective processes is likely to be poorly adapted. The more novel the body plan is, the more susceptible it will be to elimination by competition and, therefore, the more adaptive space will be required for it to become established. In short, competition tends to inhibit the establishment of higher taxa. Natural selection tends to inhibit major evolutionary change.
On The Origin of Stasis by Means of Natural Selection
Charles Darwin made the concept of major evolutionary change plausible by convincing scientists that natural selection could account for the appearance of design in nature (Horan, 1979). He would never have considered evolution to be a fact without a plausible theory of how it could occur. The very title of his book reflects the importance of an evolutionary mechanism. Although much evidence has been cited in favor of macroevolution, as it had been prior to 1859, such evidence in Darwin's own opinion would be unsatisfactory without a mechanism:
In considering the Origin of Species, it is quite conceivable that a naturalist, reflecting on the mutual affinities of organic beings, on their embryological relations, their geographical distribution, geological succession, and such other facts, might come to the conclusion that each species had not been independently created, but had descended, like varieties, from other species. Nevertheless, such a conclusion, even if well founded, would be unsatisfactory, until it could be shown how the innumerable species inhabiting this world have been modified, so as to acquire that perfection of structure and coadaptation which most justly excites our admiration (Darwin, 1859, p. 66).
In considering the origin of phyla and the stability of the major body plans, it is quite conceivable that scientists, reflecting upon the empirical evidence of the history of life on earth, might come to the conclusion that mechanisms exist which prevent major evolutionary change from occurring on a gradual step-by-step basis. Stasis is a fact of life and mechanisms must exist which account for it. Ironically, natural selection itself may play a major role in the phenomenon of higher taxon-level stasis.
p.237
Darwin was not without his critics. In his book, Darwinism: The Refutation of a Myth, Soren Lovtrup points out that "some critics turned against Darwin's teachings for religious reasons, but they were a minority; most of his opponents ... argued on a completely scientific basis." He goes on to explain:
...the reasons for rejecting Darwin's proposal were many, but first of all that many innovations cannot possibly come into existence through accumulation of many small steps, and even if they can, natural selection cannot accomplish it, because incipient and intermediate stages are not advantageous (Lovtrup, 1987).
Perhaps the most formidable of Darwin's critics was St. George Mivart. His major book, On the Genesis of Species, took aim at the notion that natural selection could account for the accumulation of the incipient stages of useful structures (Mivart, 1871). Stephen Jay Gould notes that
Darwin offered strong, if grudging, praise and took Mivart far more seriously than any other critic .... Mivart gathered, and illustrated "with admirable art and force" (Darwin's words), all objections to the theory of natural selectionó-"a formidable array" (Darwin's words again). Yet one particular theme, urged with special attention by Mivart, stood out as the centerpiece of his criticism. It remains today the primary stumbling block among thoughtful and friendly scrutinizers of Darwinism. No other criticism seems so troubling, so obviously and evidently "right" (against a Darwinian claim that seems intuitively paradoxical and improbable).
Mivart awarded this criticism a separate chapter in his book, right after the introduction. He also gave it a name, remembered ever since. He called it, "The Incompetency of 'Natural Selection' to account for the Incipient Stages of Useful Structures." If this phrase sounds like a mouthful, consider the easy translation: we can readily understand how complex and full developed structures work and owe their maintenance and preservation to natural selectionó-a wing, an eye, the resemblance of a bittern to a branch, or of an insect to a stick or a dead leaf. But how do you get from nothing to such an elaborate something, if evolution must proceed through a long sequence of intermediate stages, each favored by natural selection? You can't fly with 2% of a wing or gain much protection from an iota's similarity with a potentially concealing piece of vegetation. How, in other words, can natural selection explain these incipient stages of structures that can only be used (as we now observe them) in much more elaborated form (Gould, 1985)?
Gould goes on to point out that among the difficulties of Darwinian theory "one point stands high above the rest: the dilemma of incipient stages. Mivart identified this problem as primary and it remains so today."
There are numerous examples of organisms with systems of highly specialized, interdependent components, all of which must be integrated before they are functional and offer any selective advantage. Nature abounds in such systems of "irreducible complexity." Although there may be some examples where certain components take on some preadaptive function, these cases may be considered as exceptions to a more general rule. As a rule, any subset of the components would prove to be a burden to an organism and thus be eliminated by natural selection. Take, for example, the sensory and motor mechanism of the common bacterium, Escherichia coli, a relatively simple unicellular prokaryotic organism.
The sensory and motor mechanism consists of a number of receptors which initially detect the concentrations of a variety of chemicals. Secondary components extract information from these sensors which, in turn, is used as input to a gradient sensing mechanism. The output of this mechanism is used to drive a set of constant torque proton-powered reversible rotary motors which transfer their energy through a microscopic drive train and propel helical flagella (see Figure 7). This highly integrated system allows the bacterium to migrate at the rate of approximately ten body lengths per second.
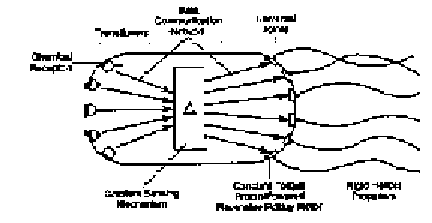 |
| Fig. 7. Conceptual block diagram of the sensory and motor mechanism of the bacterium. E. coli. |
p. 238.
Dr. Robert Macnab of Yale University concluded a major fifty page review of this mechanism with these remarks:
As a final comment, one can only marvel at the intricacy in a simple bacterium, of the total motor and sensory system which has been the subject of this review and remark that our concept of evolution by selective advantage must surely be an oversimplification. What advantage could derive, for example, from a "preflagellum" (meaning a subset of its components), and yet what is the probability of "simultaneous" development of the organelle at a level where it becomes advantageous (Macnab, 1978)?
Each component in the flagellum alone (see Figure 8) is highly specialized in its function and it is highly unlikely that many of the components, if any at all, would have any preadaptive value.
 |
|
Fig. 8. Conceptual diagram of the motor mechanism of E. Coli |
A subset of components would constitute a collection of superfluous parts which according to Darwinian theory should be eliminated by natural selection. Darwin pointed out in The Descent of Man (Darwin, 1871) that natural selection would act to preserve those individuals which were least encumbered with a superfluous part. Although this is one of the simplest organisms on our planet, it serves to illustrate how natural selection contributes to the phenomenon of stasis and helps explain the fact that most species do not arise gradually by the steady transformation of their ancestors.
Facts, Fossils, and Philosophy
Charles Darwin's classic, On the Origin of Species by Means of Natural Selection, was a persuasive and compelling argument for the idea that minor evolutionary change could be extrapolated to account for the origin of all major forms of life by purely mechanistic and materialistic processes. Writing on the origin and impact of evolutionary thought, Douglas Futuyma put it this way:
Darwin showed that material causes are a sufficient explanation not only for physical phenomena, as Descartes and Newton had shown, but also for biological phenomena with all their seeming evidence of design and purpose. By coupling undirected, purposeless variation to the blind, uncaring process of natural selection, Darwin made theological or spiritual explanations of the life processes superfluous. Together with Marx's materialistic theory of history and society and Freud's attribution of human behavior to influences over which we have little control, Darwin's theory of evolution was a crucial plank in the platform of mechanism and materialism ó- of much of science, in short ó- that has since been the stage of most Western thought (Futuyma 1986).
Were Darwin's extrapolations justified? Judging from the conclusions of many of the scientists attending one of the most important conferences in evolutionary biology in the past forty years, the answer is, probably not.
The central question of the Chicago conference was whether the mechanisms underlying microevolution can be extrapolated to explain the phenomena of macroevolution. At the risk of doing violence to the positions of some of the people at the meeting, the answer can be given as a clear, no (Lewin 1980).
Lewin's article on the conference in the AAAS journal, Science, described some of the major conflicts between the modern neo-Darwinian theory of evolution and the empirical evidence:
Evolution, according to the Modern Synthesis, moves at a stately pace, with small changes accumulating over periods of many millions of years yielding a long heritage of steadily advancing lineages as revealed in the fossil record. However, the problem is that according to most paleontologists the principle feature of individual species within the fossil record is stasis, not change...
In a generous admission Francisco Ayala, a major figure in propounding the Modern Synthesis in the United States, said, "We would not have predicted stasis from population genetics, but I am now convinced from what the paleontologists say that small changes do not accumulate" (Lewin 1980).
p. 239
If small changes do not accumulate, if stasis is the principle feature of individual species in the natural history of life, if natural selection inhibits or precludes major evolutionary change, if the order of appearance in the geologic record is systematically backwards to Darwinian predictions, and if the higher taxa are as discontinuous as they appear to be, why is it that scientists don't develop theories to explain the natural limits to biological change? The answer seems to be more philosophical than empirical. Although the question would probably lead to a more accurate description of nature, it would undoubtedly undermine the pervasive secular philosophy of mechanism and materialism which has come to dominate modern science. Although the question could certainly lead to fruitful scientific research, the thought that natural processes might actually prevent major evolutionary change from occurring would be anathema to those scientists who hold to some form of philosophical naturalism or secular religion.
In a recent AAAS conference Michael Ruse, philosopher of science and noted defender of Darwinism, admitted that evolutionary theory is based upon metaphysical assumptions.
And certainly, there's no doubt about it, that in the past, and I think also in the present, for many evolutionists, evolution has functioned as something with elements which are, let us say, akin to being a secular religion ... And it seems to me very clear that at some very basic level, evolution as a scientific theory makes a commitment to a kind of naturalism, namely, that at some level one is going to exclude miracles and these sorts of things come what may (Ruse 1993).
In other words, no matter what the evidence infers, no matter how inadequate Darwinian theory might be, only naturalistic explanations to questions of origins will be considered scientific.
If we assume science to be limited to the study
of
purely materialistic processes and we further assume that the origin of life
and the origin of the major body plans are subject to scientific investigation,
then the only truly scientific explanation of origins must be materialistic
and
naturalistic by definition.
If we assume science to be limited to the study of purely materialistic processes and we further assume that the origin of life and the origin of the major body plans are subject to scientific investigation, then the only truly scientific explanation of origins must be materialistic and naturalistic by definition. Under these rules of reasoning some form of Darwinism is the only possible "scientific" explanation of origins. Creation in any form, including theistic evolution, is by definition "unscientific."
Such reasoning is reflected in a booklet published by the National Academy of Sciences entitled Science and Creationism (1984). The influential National Academy of Sciences, representing the nation's most notable scientists, has argued that the concept of creation is not scientific:
...it fails to display the most basic characteristic of science: reliance upon naturalistic explanations. Instead, proponents of "creation-science" hold that the creation of the universe, the earth, living things, and man was accomplished through supernatural means inaccessible to human understanding (National Academy of Sciences, 1984).
For the most part, [proponents of
"creation-science" or
advocates of "intelligent design"]
have attempted to critique
evolutionary theory and to point out areas of the
theory which
are either untestable or in conflict with empirical data.
The National Academy of Sciences simply defined away all alternatives to purely naturalistic evolution by insisting that only naturalistic explanations can be considered in answering questions of ultimate origins. By definition there is no scientific alternative to the idea that
man is the result of a purposeless and natural process that did not have him in mind (Simpson, 1967).
Proponents of "creation-science" or advocates of "intelligent design" have never pretended to explain the mechanism by which the universe, the earth, living things, or man came into existence. For the most part, they have attempted to critique evolutionary theory and to point out areas of the theory which are either untestable or in conflict with empirical data. In so doing, they have inferred that purely mechanistic processes are insufficient to account for the order and complexity of the cosmos.
p. 240
This has provoked an almost religious reaction from the Academy:
"Creation-science" is thus manifestly a device designed to dilute the persuasiveness of the theory of evolution. The dualistic mode of analysis and the negative argumentation employed to accomplish this dilution is, moreover, antithetical to the scientific method (National Academy of Sciences, 1984).
Given the fact that creation events are
historical possibilities
it is entirely possible that natural processes alone
are insufficient to
account for the origin of life and all genetic information.
Berkeley law professor, Phillip E. Johnson, in his book, Darwin On Trial, concludes:
The Academy thus defined "science" in such a way that advocates of supernatural creation may neither argue for their own position nor dispute the claims of the scientific establishment. That may be one way to win an argument, but it is not satisfying to anyone who thinks it possible that God really did have something to do with creating mankind, or that some of the claims that scientists make under the heading of "evolution" may be false (Johnson, 1991).
Must scientists and science educators continue to operate under the assumption that origins must be accounted for by purely unguided naturalistic processes? Despite the fact that all other explanations of origins fail to meet the Academy's definition of science, the answer can be given as a clear "no." Given the fact that creation events are historical possibilities, it is entirely possible that natural processes alone are insufficient to account for the origin of life and all genetic information. Although philosophical naturalism requires a cosmos without a creator, science does not. Scientists working within the confines of methodological naturalism could, in fact, discover natural processes which prevent major evolutionary change from occurring, processes which explain the pervasive patterns of higher taxon-level stasis.
Without question, scientists will need to gain a much greater understanding of the processes underlying stasis. Species stasis commonly continues for millions of years, periods of time for which environmental constancy does not seem possible. Indeed, species stasis often appears to persist despite evidence for environmental change. Natural selection is, obviously, only a part of the whole picture. Internal genetic and developmental mechanisms may play an even greater role in maintaining stasis by inhibiting transitional forms from developing in the first place.
If Mivart were correct in concluding that natural selection is incompetent to account for the incipient stages of useful structures, we can only criticize him for not taking his idea far enough. He could have developed a theory of "macrostasis" and established natural selection as a key mechanism underlying the phenomenon of morphological stability and the mechanism which explains the lack of transitional forms in the fossil record. Had he done so, we might have emerged from the nineteenth century with two major theories of biological change: one accounting for minor evolutionary change and the common ancestry of the lower taxa and another accounting for the stability of the higher taxa. More importantly, we would have entered the twentieth century with theories which more accurately reflected the empirical data.
In retrospect, it seems as though Darwin was
less concerned
with the scientific question of accurately explaining the
empirical data
of natural history and more concerned with the religious or
philosophical question of explaining the design of all species without a
designer.
As we enter the twenty-first century we should pause and re-examine our presuppositions as well as our data. We must be careful not to slip into scientism and must constantly strive to most accurately describe nature, even if it means discarding some of our most cherished beliefs. As Pierre-Paul Grasse', past President of the French Academie des Sciences and editor of the 35 volume Traite' de Zoologie, expressed it:
Today our duty is to destroy the myth of evolution, considered as a simple, understood, and explained phenomenon which keeps rapidly unfolding before us. Biologists must be encouraged to think about the weaknesses and extrapolations that theoreticians put forward or lay down as established truths. The deceit is sometimes unconscious, but not always, since some people, owing to their sectarianism, purposely overlook reality and refuse to acknowledge the inadequacies and falsity of their beliefs (Grasse, 1977).
p. 241.
We must bear in mind that just because neo-Darwinian evolution is the most plausible naturalistic explanation of origins, we should not assume that it is necessarily true. Likewise, just because creation involves processes which are non-natural, we should not assume that creation events ó whether sudden or gradual ó have not occurred. It would be unreasonable to assume so. Creation events may not be subject to scientific investigation, but stasis most definitely is. "Stasis is data."
In retrospect, it seems as though Darwin was less concerned with the scientific question of accurately explaining the empirical data of natural history and more concerned with the religious or philosophical question of explaining the design of all species without a designer. Darwin's general theory of evolution may, in the final analysis, be little more than an unwarranted extrapolation from microevolution based more upon philosophy than fact. The problem is that Darwinism continues to distort twentieth century science.
©1994
References
Bowring, S.A., Grotzinger, J.P., Isachsen, C.E., Knoll, A.H., Pelechaty, S.M., Kolosov, P., "Calibrating Rates of Early Cambrian Evolution," Science, vol. 261, 3 September 1993, pp. 1293-1298
Darwin, C. (1859), The Origin of Species (Reprint of the first edition), Avenel Books, Crown Publishers, New York, 1979
Darwin, C. (1871), The Descent of Man, and Selection in Relation to Sex, J. Murray, London
Erwin, D., Valentine, J., and Sepkoski, J. (1988), "A Comparative Study of Diversification Events,"Evolution, vol. 41, p. 1183
Futuyma, D. J., (1986), Evolutionary Biology, Sinauer Associates Inc., Sunderland, MA, p. 2
Gould, S.J. (1977), "Evolution's Erratic Pace," Natural History, vol. 86, May
Gould, S. J. (1980), "Is a new and general theory of evolution emerging?" Paleobiology, 6(1), p. 120
Gould, S. J. (1985), "Not Necessarily a Wing," Natural History, October, pp. 12, 13
Gould, S. J. (1988), "A Web of Tales," Natural History, October, pp. 16-23
Gould, S. J., (1989), Wonderful Life, W. W. Norton and Company, New York, p. 49
Gould, S. J. (1991), "Opus 200," Natural History, August, p. 16
Grasse, Pierre-Paul (1977), Evolution of Living Organisms, Academic Press, New York, N.Y., p.8
Horan, P. (1979), Foreword to The Origin of Species, Charles Darwin, 1859, (Reprint of the first edition), Avenel Books, Crown Publishers, New York, p. viii
Johnson, P. (1991), Darwin on Trial, Regnery Gateway, p. 8
Kerr, R. (1993), "Evolution's Big Bang Gets Even More Explosive," Science, vol. 261, 3 September 1993, p. 1274
Levi-Setti, R. (1975), Trilobites: A Photographic Atlas, University of Chicago Press, pp. 23-45
Lewin, R. (1980), "Evolutionary Theory Under Fire," Science, vol. 210, 21 November, p. 883
Lewin, R. (1988), Science, vol. 241, 15 July, p. 291
Lovtrup, S. (1987), Darwinism: The Refutation of a Myth, Croom Helm Ltd., Beckingham, Kent, p. 275
Macnab, R. (1978), "Bacterial Mobility and Chemotaxis: The Molecular Biology of a Behavioral System," CRC Critical Reviews in Biochemistry, vol. 5, issue 4, December, pp. 291-341
Mivart, G. (1871), On the Genesis of Species, London, Macmillan
National Academy of Sciences (1984), Science and Creationism: A View from the National Academy of Sciences
Raup, D. (1979), "Conflicts Between Darwin and Paleontology," Field Museum of Natural History Bulletin, vol. 50 (1), p. 24, 25
Raup, D. M. (1981), "Evolution and the Fossil Record," Science, 17 July, p. 289
Ruse, M. (1993), "Nonliteralist Antievolution," AAAS Symposium "The New Antievolutionism," February 13, 1993, Boston, MA
Simpson, G. G. (1944), Tempo and Mode in Evolution, Columbia University Press, New York, p. 105, 107
Simpson, G. G. (1967), The Meaning of Evolution, p. 345 (revised edition)
Thompson, W. R. (1956), Introduction to The Origin of Species, (Reprint of the first edition), Charles Darwin, Everyman Library, no. 811, Dent
Valentine, J.W., Awramik, S.M., Signor, P.W., and Sadler, P.M. (1991), "The Biological Explosion at the Precambrian-Cambrian Boundary," Evolutionary Biology, Vol. 25, Max K. Hecht, editor, Plenum Press, New York and London, p.284
Valentine, J., and Erwin, D. (1985) "Interpreting Great Developmental Experiments: The Fossil Record" Development as an Evolutionary Process Rudolf A. Raff and Elizabeth C. Raff, Editors Alan R. Liss, Inc., New York, pp. 71, 95, 96
Wise, K. (1991), "Changing Stasis," Origins Research, vol. 13, no. 1, p. 20
