Science
in Christian Perspective
Science
in Christian PerspectiveScience
in Christian Perspective
Roy M. Allen, Sc.D.
368 Ridgewood Ave.
Glen Ridge, N.J.
From: JASA, 2,
(June1950): 18-25.
Figs.
For establishing an impartial concept of mechanistic evolution versus special
creation no other organ or structure of animate life offers such positive information
as the organ of vision. Depending on the viewpoint, its evidence can be interpreted
in three radically different ways.
First, there is the position of the evolutionist who sees in eye structures,
when arranged in an ascending scale of complexity, abundant, justification for his
belief. Then there is the more logical appreciation of the failure of eye structures as a whole to fit into any possible taxonomic relationship which evolution
can devise. Complete nullification of any progressive arrangement mechanistic
development offers results from comparison of numerous structures occurring throughout the
gamut of animal life. No plausible explanation of many of these structures,
apart from controlled design on the part of a Creator will suffice. Lastly, there
is the highly developed human-eye with its marvelous mechanical and optical perfections testifying to an unbiased mind the absolute necessity for every part to have
been planned by a Master Designer with skill and knowledge far surpassing the utmost
the human mind can conceive.
The usual procedure in formulating evolutionary theories, of whatsoever sort
they may be, is largely consideration of anatomical structures as a whole, without
regard to radical variations in component parts. Progressive development of various
organs, structures, etc., must be assumed to follow as a matter of course. Failure
of any of these to keep pace with & general development scheme is not considered subversive to an evolutionary concept, even-when no suitable theory can be offered to
account for conditions as found. As a result of this position evolution feels justified in taking such a structure as the eye and by proper arrangement of various forms.,
tracean assumed development from origin all the way through to perfected organ4 Such
demonstration frequently carries conviction as to the correctness of the evolutionary
concept to minds not skilled in the analysis of all the factors involved. Because of
this, it appears advisable first to examine the train of reasoning adopted by mechanistic evolution in the cas.e of the eye before presenting facts which cannot be made
to fit into a mechanistic mold.
In the Amoeba there exists no evidence of light receptive differentiation, the
entire cell functioning to perceive variations in light intensities. However, this
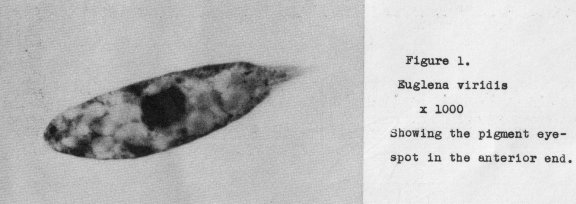
is not the case with all unicellular organisms* In Euglena. the first evidence of a
localized light percepter is found in an anterior pigment spot. The Euglena pictured
in Figure 1. at a magnification of 1000 diameters, shows this early stage of what evolution assumes is to develop
through the ages of time into an eye. Certain it is
that light reception, when arriving at a more evident stage, is associated with an
essential pigment foundation, such as present in Euglena.
Progressing to the metazoa we find certain cells functioning as light-sensitive
detectors, occasionally without pigment (as for example, in the Hydra, Lumbricus, etc;)
but here a new departure in structure, also essential to a final eye form, is
introduced. The light-sensitive cells are associated with a nerve plexus or more highly
developed nervous system* However, we do not have to go far in the usual types of
metazoan life to see both the pigment foundation and the specialized light-sensitive
cells combined in function and associated with a nerve system.
The rotifera constitute a good example of this. Figure 2 shows a head-on view
of a minute rotifer, Pedalion, at a magnification of 400 diameters, in which primitive
eyes are evident. These consist of a pigment spot at the end of a terminal nerve cell
forming part of a nerve plexus. But in spite of the minute size of this rotifer (only
250 microns In greatest dimension), a new principle in eye structure appears.
The eyes
are paired and spaced some distance apart, While different species of rotifers vary
in the number of eyes present, - from none, to several pairs - the common case is one
pair. In the extremely primitive eyes present in Pedalion, it appears that the photoreceptor is the pigment granule which may function through a heat-absorbing effect,
detected by the associated nerve cell and made evident to the entire body via the
nerve plexus. It is possible that with paired detectors a directional faculty is
present.
Further specialization in other forms of life amplifies the pigment into multitudinous pigment cells arranged in the form of a hollow cup; also the single nerve
cell has become many situated within the pigment sector. Illustrative of this stage
is the Leech (Glossiphonia) the eyes of which appear in a cleared and mounted specimen as in Figure 3 (magnification 300x). Other instances of this stage occur in widely
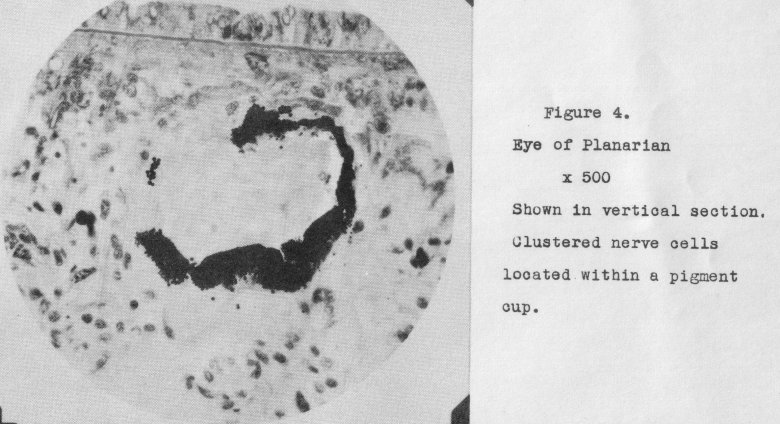
different species. The planarians offer one such example. A vertical section through a Planarian eye is shown in Figure 4, at 506 diameters. Thus far there
does not appear any uniform arrangement of the light-sensitive nerve cells; they merely fill the pigment cup.
The relative size of an animal is no criterion of the degree of development of
the organ of vision, nor, as will be soon later, is the degree of development in the
scale of life a safe guide. Up to this point the eyes are elementary or strictly
primitive. No image can be perceived, except possibly as an opaque object comes between the eye and the light
source, thus casting a shadow, For the formation of an
image a lens (or its equivalent) is essential. The inception of such lens can be
represented by the eye of a minute water-mite (Hydracharina) as seen in Figure 5
(magnified 450x). Here we see a nearly spherical transparent lens placed in front
of the nerve cells and projecting within the rim of the pigment cup. The entire eye
is a unit, although as in the case of the more elementary form, still located within
the body.
While we are not, at this time discussing compound eyes, mention can be made of
the equivalent stage to be found in some of the entomostraca (e.g., Daphnia) whore
several unit eyes with lenses are grouped together to form a clustered eye, looking
very much like a miniature raspberry. A picture of this eye will be shown later.
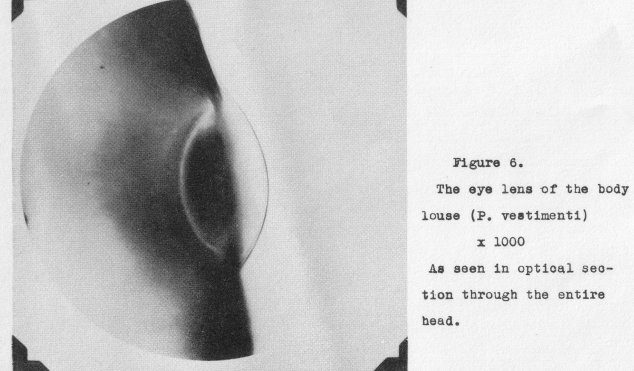
The perfected lens of the simple eye can be illustrated by that of the body
louse (P. vostimonti) where it is no longer within the body, but formed by a lentioular portion of transparent
chitin. This lens, which is still minute, appears in optical section at a magnification of 1000 diameters as in Figure 6. 7ollowing the advent of such a lens and improvements in its design we find corresponding changes required in the photo-receptor cells. To be efficient in the detection of the image
formed by the lens these can no longer be haphazardly placed within the pigment cup.
They must be vastly increased in number, reduced in diameter so as to cover the least
possible area of the image, and be disposed in the focal plane
of
the lens.
At this stage of
developed eye structure, especially characteristic of the invertebrata we have many manifestations of the form the organ of vision may
take.
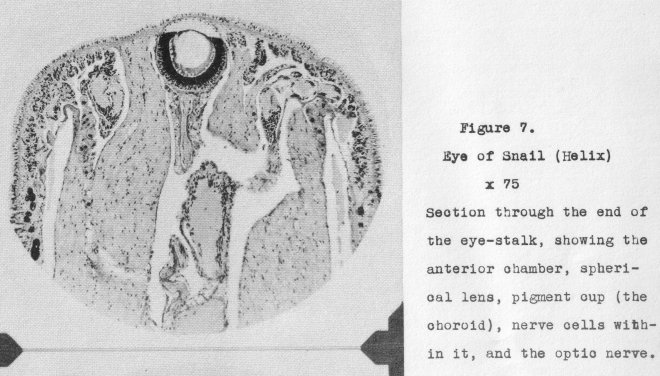
Illustrative of them may be cited that of the snail (Helix) which is shown in Figure 7
at a magnification of 75x, the individual eyes (ocelli) of insects, the eyes of spiders,
those of the scallop (Pecten), and the ommatidia of compound eyes. Basically, however,
these all possess certain structures in common. 5uch are: a corneal membrane, a crystalline
(or, cellular) lens, light-sensitive retinal cells (rods) systematically
disposed in the focal plane of the lens, an enclosing pigment layer, and an optic nerve
connecting the retinal cells to the brain or central nerve ganglion. In addition,
this stage of developed eye structure, especially characteristic of the invertebrata
where there may be found in some instances an anterior chamber between the cornea and. the
lens, migrating pigment for adapting the eye sensitivity to variations in light intensity, and other minor structures having specific functions.
Thus step by step, evolution envisages progress in the development of the eye through the invertebrata, with
ever increasing complexity of structure and refinement
of operation until it reaches a high degree of function in the vertebrates, finding
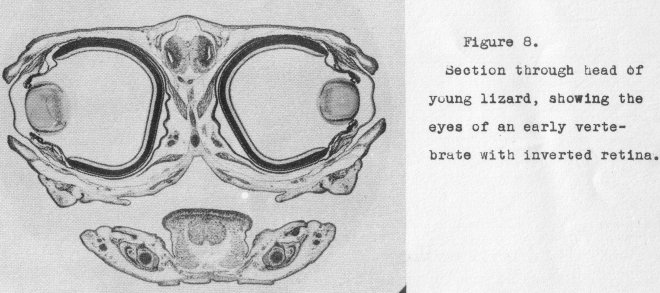
its full development in the mammals and man. Figure 8 shows in a section through the
head the stage reached in the reptilia. Eyelids, anterior chamber, iris, and primitive muscle control are evident. It is but a step more to
the final perfected eye of
the mammals and man.
How well the eye, when viewed in this light, appears to confirm the claims of
evolution! It is not until we examine some of the evidence not conforming to the
claims of evolution we discover that mechanistic evolution., regardless of the particular theory applied, does not, and cannot, explain how the eye came into being.
Let us take the case of Amphioxus in the scale of life assigned to it by evolution. It is ranked among
the most primitive of the chordata since it possesses the
beginning of a back-bone. Presumably, therefore, it has evolved above or beyond the
invertebrata, forming a connecting link with the fish, via the Lampreys (Petromyzon,
etc.) The general anatomical relationships would seem to bear this out. But just
where in the evolutionary tree did it branch off? If its position were to be established on the basis of eye structure
amphioxus must be placed among the lowest forms
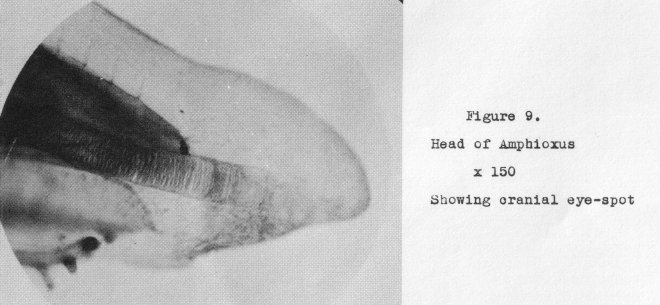
of invertebrate life. The mass of pigment usually designated the cranial eye-spot
(shown in Figure 9 at a magnification of 150 diameters) is little more than the pigment spot in the
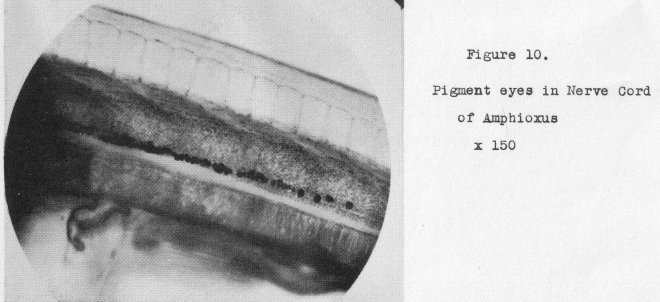
ouglena. This, however, is not the only eye amphioxus possesses, Scattered throughout the anterior portion of
the nerve cord are numerous other eyespots (Figure 10, also 150 diameters)
consisting of two cells each-- a pigment cup
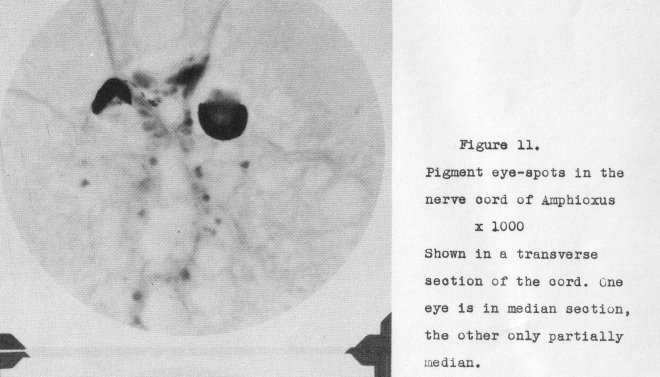
and a light-receptor cell lying within it, A couple of these are shown in a
transverse section in Figurc. 11, at a magnification of 1000 diameters. What an apology
for an eye, or eyes, in an animal placed at the beginning of the backbone
animals!
The larval eyes of the related Tunicates, lost completely in the adult stage, are, in
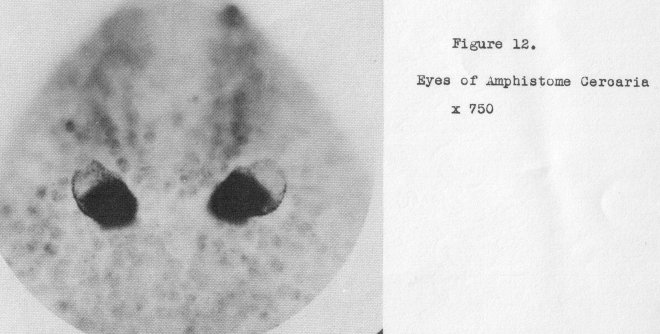
the same, category. As a matter of fact, the eyes of some amphistome, cercariae, also
lost in the adult stage, are higher in the scale than amphioxus, as is evident from
Figure 12 (at 750x) where they are seen to be paired. The question as to why an animal so
highly developed as amphioxus in other respects does not at least possess paired eyes, is unanswerable from an evolutionary standpoint.
Among other equally unanswerable questions (which are multitudinous)related to
eye structure may be cited the following:
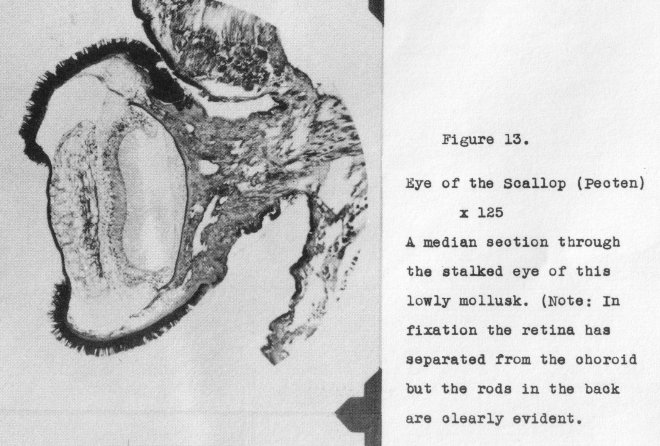
Why does the lowly mollusk Pecten, possess a hundred or more individual stalked
eyes
each with a well-formed cellular lens and an inverted
(vertebrate), type of
retina? This eye is shown in section
in Figure 13, at a magnification of 125 diameters.
How to explain the occurrence of stalked eyes in such widely diversified forms of
life as Pecten, Helix, Squilla, Cambarus and, most remarkable of all, the African fly,
Diopsis, shown in Figure 14? Where is the common starting point? are some of
these stalked eyes simple and others compound?
How to account for compound eyes? True, it is possible to find connecting forms
suggesting a possible
link;
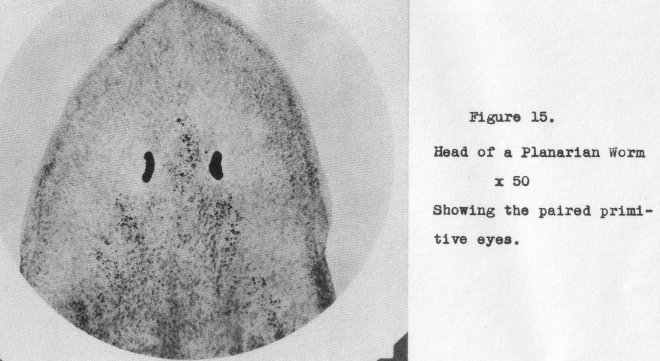
of development when eyes only are considered. For instance, in Planaria we have
the common paired primitive eye (Figure 15, at 50 x)
while in the closely related form Polycalis (Figure 16, at 60 x) a multitude of
primitive eyes, more or less uniformly scattered throughout the anterior region
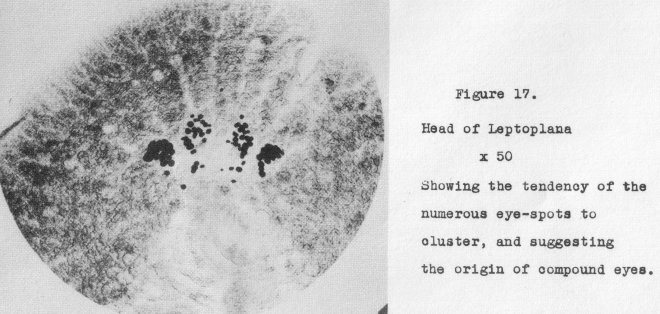
take the place of the paired eyes. Going now to Leptoplana, another near relative,
the considerable number of eyes have become clustered (Figure 17, at 50x). From
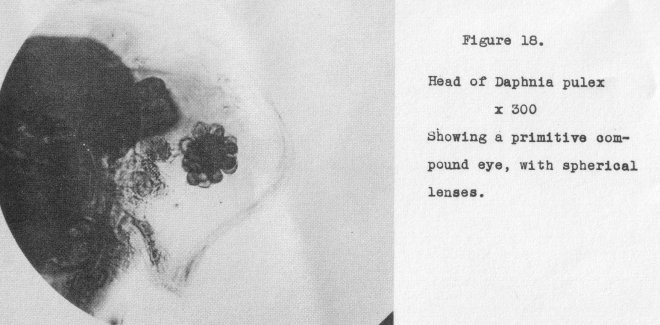
here we might progress to a primitive compound eye such as found in Daphnia, Figure
18, at 300x, and so on through various stages until we roach the highly developed
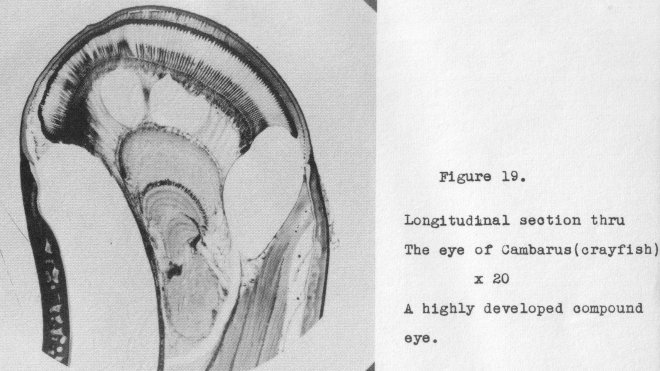
compound eye of the crayfish shown in section in Figure 19, at 20x. But are we to
reason from this arrangement that each organism in the chain is an ancestor of those
showing progressive advance toward the highly developed compound eye? Even evolutionary biologists cannot so claim; then how construct a genealogical tree which will
satisfy eye structure?
How to explain the fact that some insects possess simple paired eyes, others
have
only compound eyes while many have both, the common case in this latter instance
being
the presence of three triangularly placed simple eyes (ocelli) between the compound
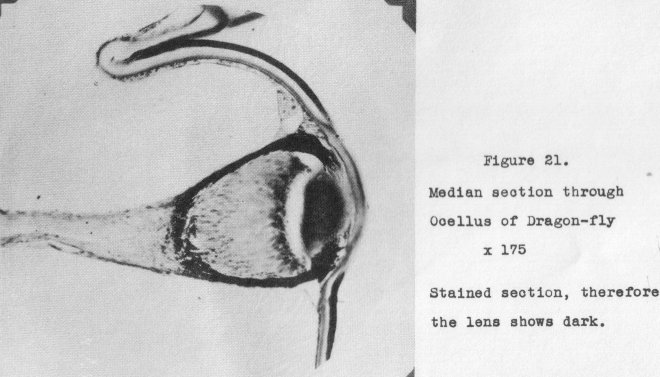
eyes? Figure 20 shows a section through the head of a dragon-fly, median through two
of the ocelli as well as the compound eyes. The structure of the ocelli is
brought
out in Figure 21, magnified 175 diameters. (Note: this is from a stained section,
hence the lens shows dark.)
Why are the spiders and their close associates usually provided with simple paired eyes, from one to
several pairs? It would seem more in keeping with their place
in taxonomic classification for them to have compound eyes. More curious yet, why
are the anterior median eyes of the inverted, or prebaccillary type corresponding
in general to vertebrate eyes in development and design, while all the other pairs
are of the erect or probacillary type? Here is something, for evolution to ponder over.
Then again, why are some spiders (the nocturnal species) provided with a tapetum?
In regard to the presence of a tapetal layer back of the retina, how can evolution account for its existence in
the various forms of life in which it occurs? We
find it in some spiders, some insects, some birds (e.g., the owls), bats, the cat
tribe and other widely separated forms. Since the tapatum enhances vision in weak
light and thus becomes an additional refinement, why have not all the mammals and
man been the recipient of such ultra refinement? Why are there so many different
types of tapetal structure, all serving the same purpose?
If there were no other evidence against a mechanistic evolutionary concept, except the case of the eye of Sepia we would still have ample to justify repudiating
it absolutely. As previously intimated, if evolution of all forms of life from some
original form was brought about by purely external mechanistic forces apart from controlled origin by a Creator, then each type of organism must have branched off from
pre-existing species at some stated time and place. In other words,, the construction
of a genealogical tree becomes not only possible, but necessary. Each new development must have been built upon what had preceded. Lot us now raise the question,
"Where did the mollusca, which includes the Sepia, leave the stem which later produced the vertebrates?" Certainly this must have occurred while the eye development
was still very primitive. Then how can we explain th6 parallel development of the
eye of Sepia and the mammalian eye to a point of substantially equal efficiency on
the one hand, yet one a strictly invertebrate eye while the other is the typical
inverted eve with its retinal origin via the neural tissue.
Few biologists appreciate the remarkable refinements found in the eye of Sepia
and the degree to which it approximates the mammalian eye. Figure 22 shows a median
section through the head and eyes of Sepia. In it can be seen the eyelids, corneal
membrane, iris,, the remarkable two-piece lens providing correction for aberrations
in a manner similar to the combined anterior chamber of the human eye, the
suspensory ligaments., the pigmented ciliary processes, the retina, the choroid.. the sclerotic
shell and muscles for moving the eye. How could this eye have developed independently so as to approximate the
mammalian eye from the point in past ancestry from which
they both emanated?
How can we fit into an evolutionary scheme the pin-hole camera eye of the Pearly
Nautilis? Here there is no lens, the sea-wateor filling the eye cavity and impinging
directly on the retina, the image being formed by the minute opening in the
iris.
And what shall we say about the pineal eye----so obviously a primitive eye structure in many of the lizards,, even
to the extent of functionability in Hatteria, yet an interior endocrine gland in
mammals?
These are but a few of the many problems presented by eye structure which cannot be solved on a mechanistic
evolutionary basis. They all testify to design in
created life, completely under control of a Master Designer.
This brings us to a final discussion of evidence gleaned from the eye as to
extent to which the design of the eye has been perfected by Him who doeth all
things
well. Since many of the facts already presented demonstrate this as regards the
lower
orders of life, we can now confine ourselves largely to consideration of the perfected
eye given to us humans.
A paper on "The Eye as an Optical Instrument" by Dr. Frank Allen was presented
at the 1948 Convention of the Association. In this was outlined the marvelous performance of the
human eye from the optical standpoint. It will therefore not be necessary to review this phase of the subject at this time, but
rather point out some of
the mechanical and physical features which could not have been the result of chances
That some of the intricacies of the eye present problems not explainable by any theories
of mechanistic evolution has been tacitly admitted by numerous evolutionists. For
example, witness Darwin's own statement: "To suppose that the eye with all its
inimitable contrivances for adjusting the focus to different distances, for admitting different amounts of light, and for the correction of spherical and chromatic aberration,
could have been formed by natural selection, seems, I freely confess, absurd in the
highest degree." However, he goes on to say, "Reason tells me that if numerous gradations from a simple and imperfect eye to
one complex and perfect can be shown to exist, each grade being useful to its possessor, as is certainly the case; if further,
the eye over varies and the variations be inherited, as is likewise certainly
the case;
and if such variations should be useful to any animal under changing conditions of
life, then the, difficulty of believing that a perfect and complex eye could be formed
by natural selection, though insuperable to our imagination, should not be considered
as subversive of the theory," (Origin of Species, Chapter 6) In other words, in spite
of the absurdity and the impossibility of even imagining it to be true, if you are going to accept the theory you must also accept the absurdity: While he is limiting
these remarks to his own pot theory, they will be found applicable to all others as
well.
Recognizing that the eye functions as a camera, it is not surprising that the
outer spherical box consists of a firm, hard shell (the sclerotic) capable of holding
all parts in rigid relation to each other, This is coated on the inside with an
opaque black layer (the choroid) against which the sensitive film (the retina) is
located, The relationship of these three layers is seen in section in Figure 230
The front of the camera is taken up with the compound lens (the anterior chamber and
crystalline lens) combined with its between the lens' diaphragn (the iris), and shutter (the eyelids), Automatic focusing and light intensity control are provided, far
beyond anything possible in the highest grade cameras today. All of this suggests careful design by One cognizant of all the, physical laws involved.
Just as in a camera, the best definition occurs in the area surrounding the optic
axis, so we find in this area of the eye a central depression, the fovoa, where vision
is of the highest order and color perception at a maximum. Figure 24 shows a section
through this minute pit and the optic nerve, A higher power detail of the fovea is
seen in Figure 25. When we wish to see anything clearly we must bring it to focus in
the fovea. Because of this
the most intense and least refracted light falls on this
area. To off-set this, special provision is made to prevent undue damage to the retina surrounding the fovea, especially by short waves, This is the incorporation of
a yellow filter in this area, known as the macula lutea (yellow-spot). Thus we have
the camera analogy carried still further. All dame ns are aware of the purpose
and value of a yellow filter in this same connection.
The light receptor cells of the retina are the rods and cones. Both pick up
light rays but only the cones are able to differentiate various wave frequencies(6olor).
At the fovea there are no rods present but as the distance from the fovea increases
the rods become more numerous until at the periphery there are no cones. Since the
light falling on the outer zones distant from the fovea is much less, no yellow filter
is required but provision must be made to equalize the lesser light intensity. A
supersonsitizing is required. Since the eye sensitivity is at a maximum in the middle
of the spectrum,, gradually falling to zero at the blue and red ends, those end colors
must be enhanced to register. Some years ago it was discovered that camera film could
be made more sensitive to any desired color by the incorporation of a dye of that color
in the
film. At that time it was not known that the Maker of the eye employed this
same principle to enhance the color perception at the two ends of the spectrum by the
presence of a blue-red (purple) dye in the retina. This is known as rhodopsin, or
'visual purple'. Can anyone conceive of this being brought about by blind
chance, as
required by mechanistic evolution?
A complete discussion of all the other marvelous adaptations found in the eye
would extend the length of this paper unduly; therefore only brief mention can be made
of most of theme
The entrance of the optic nerve through the retina necessitates a blind spot in
each eye, That this blind spot will not cause us any inconvenience, it has been arranged so that the optic nerve
enters each eye at an interior angle and thus the blind
spots cannot coincide. Thus we are not cognizant of it when both eyes are in
use.
The function of the choroid layer back of
the retina is to absorbs such light as
passes through the retina and therefore not utilized. The pigment is extra thick surrounding the blood vessels since if all the light were not absorbed, as is the case
when extremely brilliant light enters the eye, the red reflection caused by the blood
would make us see red! Similarly, there are no blood vessels in the cornea which would
act as a red filter.
In order to perceive fine detail--up to over 200 lines per inch the rods and
cones must be of extremely fine diameter and positioned so that the light strikes them
on the ends. There are about 2,000,000 rods and cones in each eye. Everyone of these
must be able to transmit its own impulse to the brain via the optic nerve, which can
be likened to a telephone cable serving two million phones.
Since there is a reflecting surface on the retina and the choroid cannot absorb
100% of the light striking it, some light must be reflected back toward the
front.
Such as strikes the Ions passes outside the eye and does no harm, but the reflection
being general, some light impinges on the entire back surface of the anterior chamber#
Should this be re-reflected toward the retina it would fog the image and poor vision
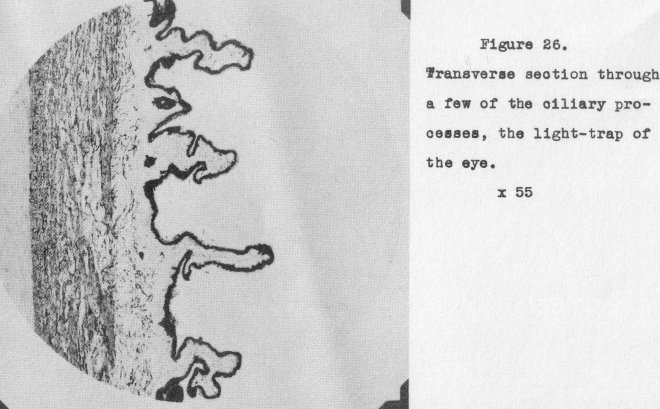
would result. To prevent this occurring, the back surface surrounding the lens is
provided with a light trap known as the ciliary processes. Those are pigmented
corrugations circularly disposed about the lens. A cross-sectional view of a few of these is
shown in Figure 26 (magnified 55x). It is obvious that any light striking such a
surface as these present will be securely trapped within the processes themselves.
The marvelous correction for optical aberrations provided by the anterior chamber, the aspheric form of the cornea and crystalline
lens and the varying refractive indices of the latter cannot have been the result of blind chance or the operation of
any external conditions (e.g., natural selection, use or disuse, environment, isolation,
or other) as predicated by various theories of mechanistic evolution. They
testify to
preconceived original design on the part of a Creator. The same can be said of the
complete automatic co-ordination of every portion and function of the eye-muscular
movement in every direction for bringing the
image
of an object to the fovea, focusing for distance (anything from a
few inches to infinity), control of light intensity
by the iris diaphragm, independent movement of the eyes to provide binocular vision,
automatic lubrication of the eyelids, and their almost instantaneous closing in the
event of threatened danger to the eyes. Mention might also be made of the provision
for controlling the amount of lubrication by means of the tear ducts.
One feature of design in the crystalline lens not so commonly known is the nature of the fibers constituting the interior portion of the lens. Since these fibers
are positioned in a parallel manner it is obvious a striation effect would result and
vision be impaired accordingly. How did the 111aster Designer solve this problem? By
the simple oxpedient of providing irregular serrations on the edges of the fibers so
that as they fit together no multiplicity of straight surfaces can occur. This can
only be observed by separating out the individual fibers, then the serrations become
evident., as can be soon in Figure 27. Mechanistic evolution cannot account for this
provision.
Perhaps no function with which the eyes are endowed is so spectacular as light
detection itself, especially color perception. Light waves traveling at 1860000 miles
per second and vibrating anywhere between 450 trillion and 750 trillion per second enter the eye and are picked up by a
set of cells especially tuned to receive them.
The stimulus set up in those cells must be converted into vibrational energy impulses
of an entirely different sort for transmission over the nerve cables to the central
station, the brain. This converted energy does not travel at 186,000 miles per second but only a few hundred feet per second, yet it must faithfully reproduce the variations in light intensities and
frequencies. The retina itself is a very complicated
structure, consisting of ten separate layers, only one of which is the rod and cone
layer. The transformation of light energy into nerve energy is accomplished in these
various other layers, that is, they constitute a two-stage step-down transformer, as
it were. Since each rod or cone may be transmitting a signal different from all others,
the transformation is a complicated process.
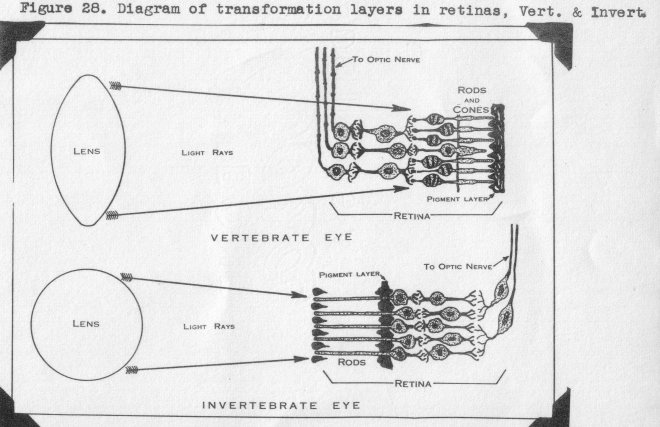
In other portions of the nerve us system throughout the entire body, terminal
nerves are continuous through ganglia and various types of nerve fibers., and thus are
connected directly with the brain. Not so the rod and cone cells with their nuclear
portion. Minute fibrils from the latter terminate in a reticular layer, Likewise the
next nuclear layer has fibrils terminating in this layer without, however, being in
any way directly connected to the rod and cone cells. This same construction is repeated in a second reticular layer, into which ganglion cell fibrils also enter, The
ganglion cells are connected via the optic nerve to the brain. Figure 28 shows this
structure diagramatically, both for vertebrate eyes and highly developed invertebrate
eyes. The transformation of light energy to nerve energy takes place in these reticular layers, a two-stage process. The various layers in the human retina are shown in
section in Figure 29; the retina 'of Sepia, an invertebrate eye where the same condition obtains, is shown in Figure
30. Could uncontrolled evolution have ever produced
this remarkable functioning structure, though we grant it a hundred billion years to
,bring it about? But further, on the same subject, a remarkable confirmation; even
the compound eyes of insects, with their thousands of ommatidia must function in
a
similar manner in order to transform light energy to nerve energy! Study of the
various layers occurring in the eye of the common house-fly, as revealed in Figure
31 will confirm this.
If the rank and file of individuals were asked why we are provided with two eyes,
the common reply would be that if one were damaged or destroyed we would still have
another to fall back on. This, however, is but one advantage; the main purpose is to
provide binocular vision. The two eyes see any object from different angles, the muscular control automatically moving the eyes independently to focus the object
on the
fovea of each, Then the, brain automatically measures the parallax angle between the
eyes and computes the distance away. Also it computes at the same time the focal
length of the lens required to bring the object to a correct focus, adds those two
values together and true stereoscopic vision results. Truly this is a remarkable
design.
In this connection mention should be made of the eyes of birds, so situated on
opposite sides of the head that true binocular vision is impossible. Yet birds fly
fast and must be able to judge distances accurately. How are they provided for in
this respect? The method is unique. The eyes arc equipped with two fovea, located
a little distance apart.. Vision is perfect in -either fovea, hence by the
simple expedient of moving the head through a small angle an object can be brought to focus
first on one fovca, thon on the other and the angle through which the head moves tells
the bird just what our binocular vision does. It is interesting to observe this head
movement in a canary or chicken as they examine a visitor closely.
There are many other eye structures equally positive in their testimony to a Master Designer back of each and
every detail, That this applies to all animate creation as well can be illustrated by a single instance. Spiders shed their chitinous
skins periodically and the eye lens of the spider is integral with the skin, Therefore, when the skin goes, the lens goes with it. But back of the lens is a special
layer of cells, the sole function of which is to provide a now lens whenever required,
This rejuvenating layer is soon in a vertical section through the eye lens of a
spider
in Figure 32. Even the lowly forms of life have a God who is able and adequate to supply every
need.
In closing, let it be pointed out that to those of us who believe the inspired
Word and know a God able to 1;ring things to pass, the issue is not whether all
physical life is one in essential being, since God could have created it in exactly that
way if He chose,. The issue on which we stand is that He conceived, planned and executed every form of life according to the counsel of His own will; therefore,
mechanistic evolution, as expounded under many and devious theories had nothing whatever to do
with it.